

Presenilin 1 Maintains Lysosomal Ca(2+) Homeostasis via TRPML1 by Regulating vATPase-Mediated Lysosome Acidification
- PMID: 26299959
- PMCID: PMC4558203
- DOI: 10.1016/j.celrep.2015.07.050
Presenilin 1 Maintains Lysosomal Ca(2+) Homeostasis via TRPML1 by Regulating vATPase-Mediated Lysosome Acidification
Abstract
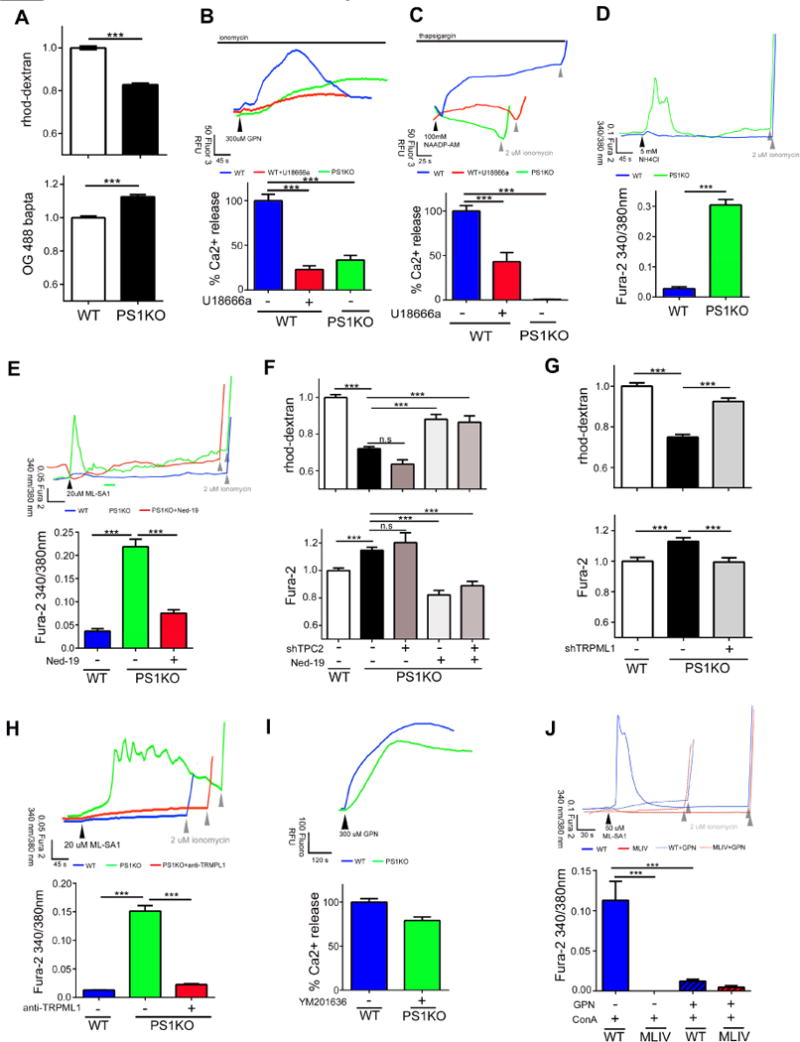
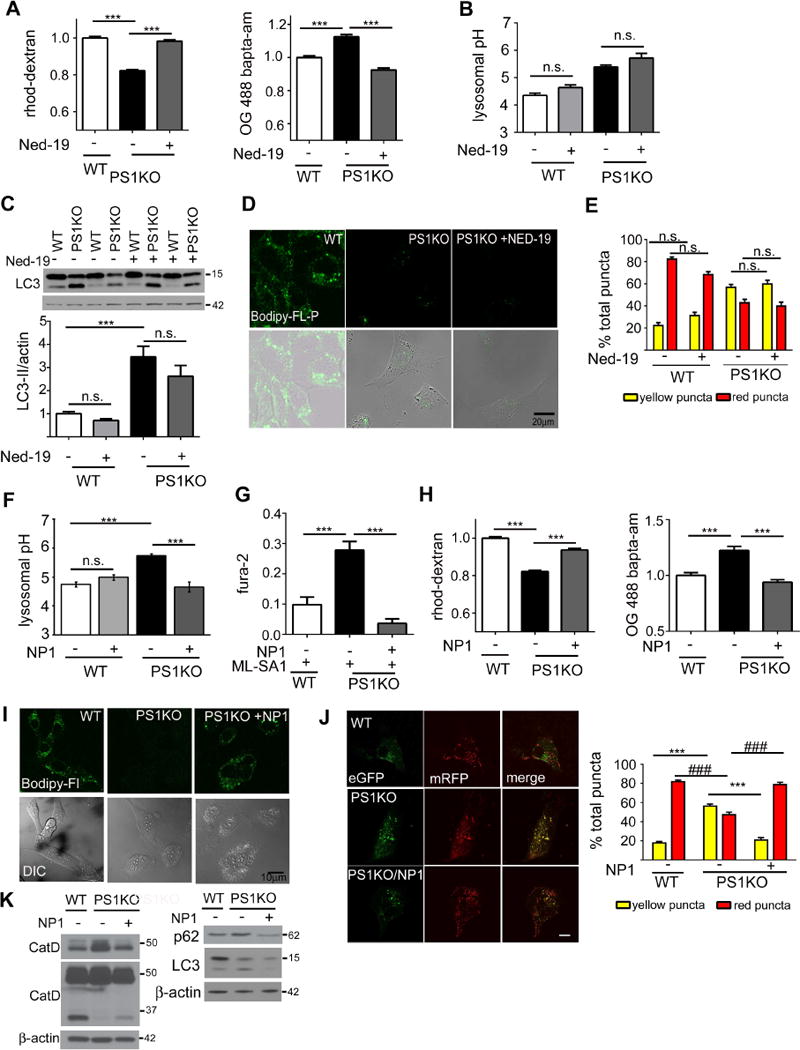
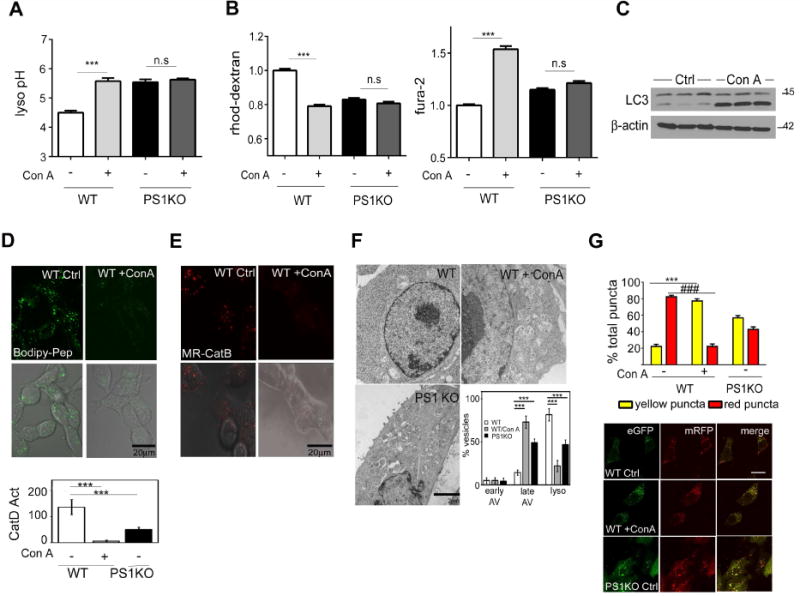
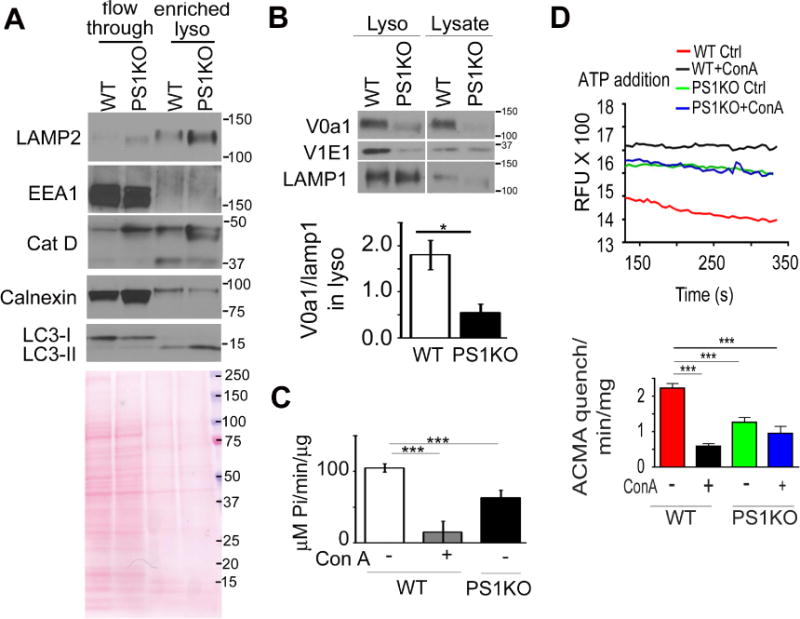
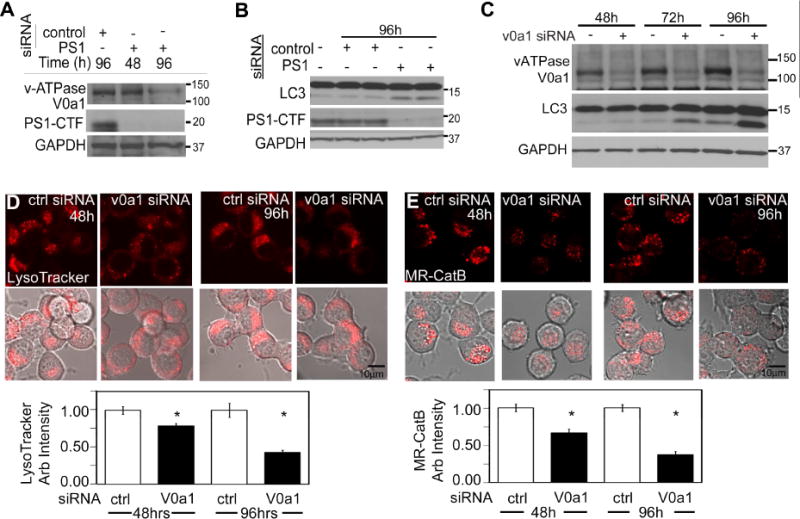
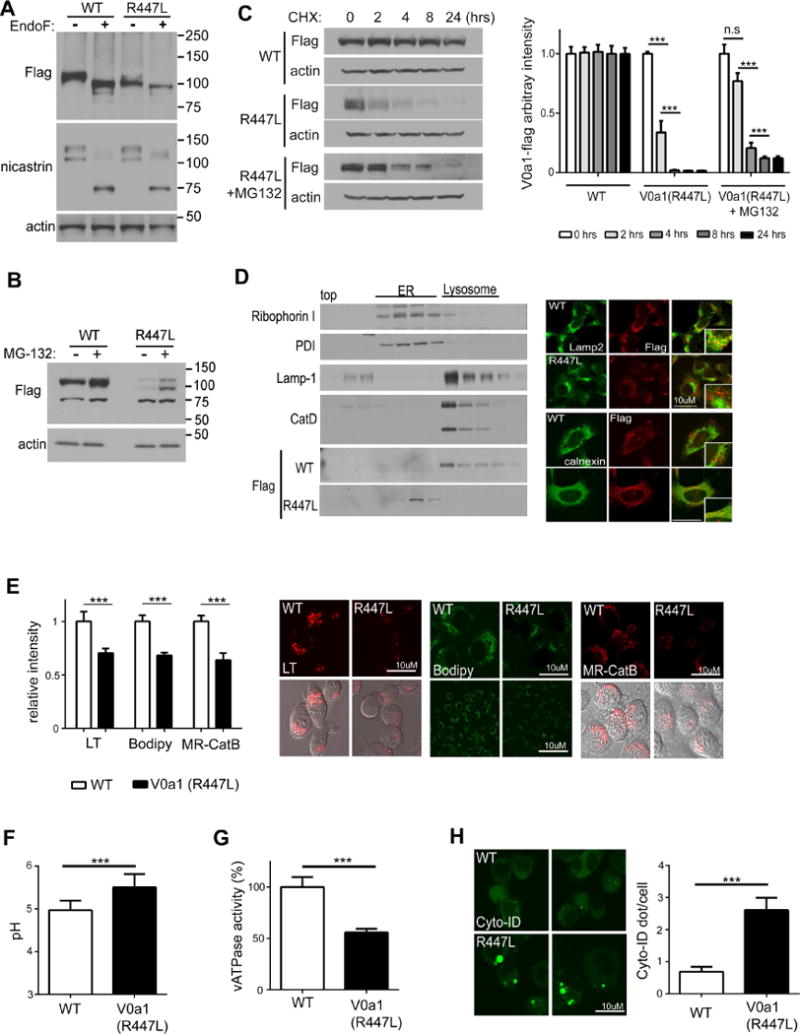
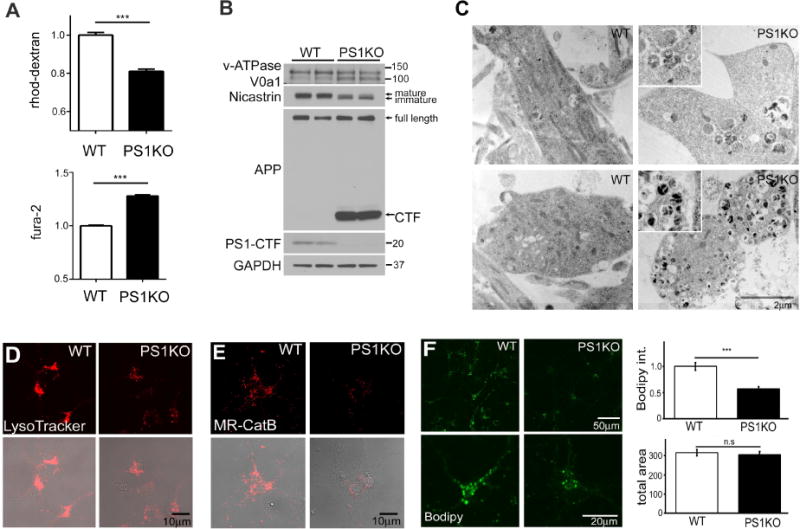
Presenilin 1 (PS1) deletion or Alzheimer's disease (AD)-linked mutations disrupt lysosomal acidification and proteolysis, which inhibits autophagy. Here, we establish that this phenotype stems from impaired glycosylation and instability of vATPase V0a1 subunit, causing deficient lysosomal vATPase assembly and function. We further demonstrate that elevated lysosomal pH in Presenilin 1 knockout (PS1KO) cells induces abnormal Ca(2+) efflux from lysosomes mediated by TRPML1 and elevates cytosolic Ca(2+). In WT cells, blocking vATPase activity or knockdown of either PS1 or the V0a1 subunit of vATPase reproduces all of these abnormalities. Normalizing lysosomal pH in PS1KO cells using acidic nanoparticles restores normal lysosomal proteolysis, autophagy, and Ca(2+) homeostasis, but correcting lysosomal Ca(2+) deficits alone neither re-acidifies lysosomes nor reverses proteolytic and autophagic deficits. Our results indicate that vATPase deficiency in PS1 loss-of-function states causes lysosomal/autophagy deficits and contributes to abnormal cellular Ca(2+) homeostasis, thus linking two AD-related pathogenic processes through a common molecular mechanism.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Avrahami L, Farfara D, Shaham-Kol M, Vassar R, Frenkel D, Eldar-Finkelman H. Inhibition of glycogen synthase kinase-3 ameliorates beta-amyloid pathology and restores lysosomal acidification and mammalian target of rapamycin activity in the Alzheimer disease mouse model: in vivo and in vitro studies. J Biol Chem. 2013;288:1295–1306. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
