

Lack of Evidence for PKM2 Protein Kinase Activity
- PMID: 26300261
- PMCID: PMC4548833
- DOI: 10.1016/j.molcel.2015.07.013
Lack of Evidence for PKM2 Protein Kinase Activity
Abstract
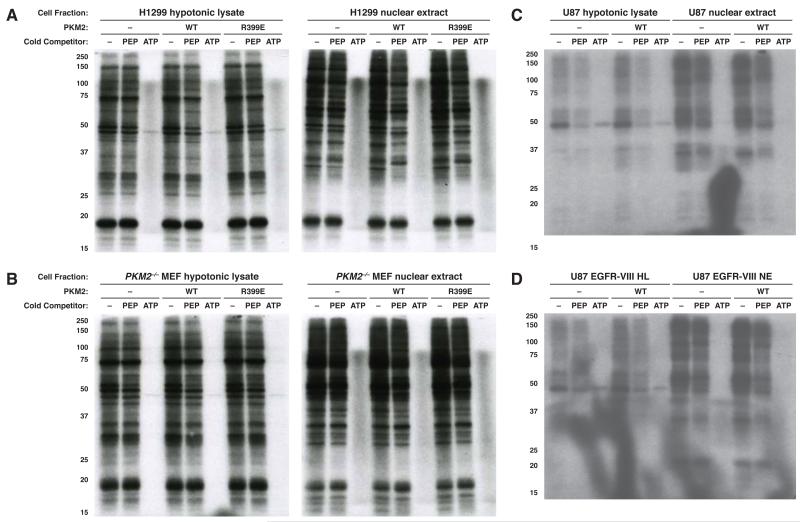
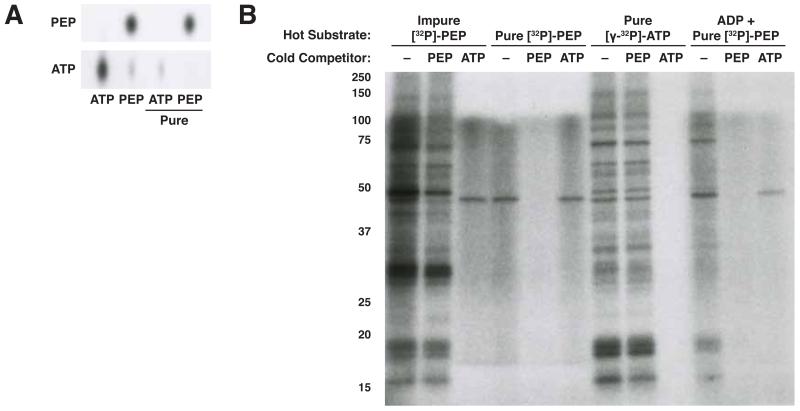
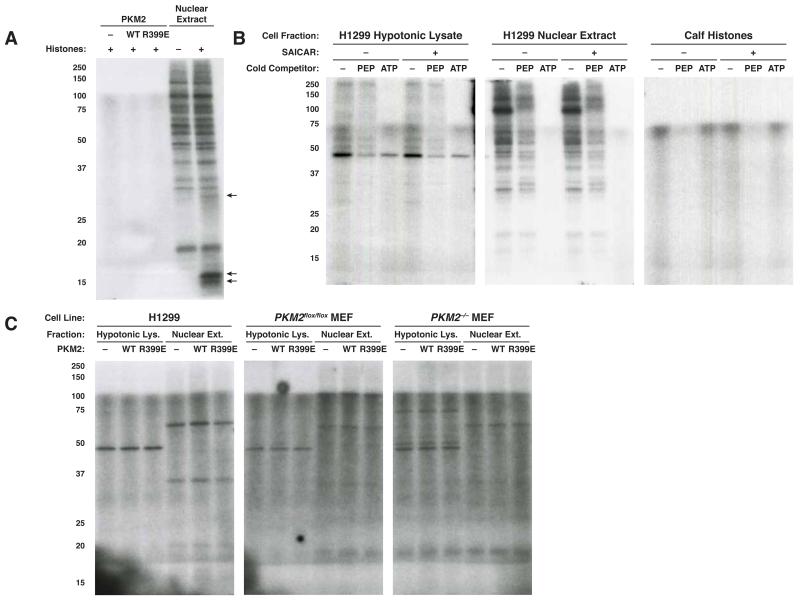
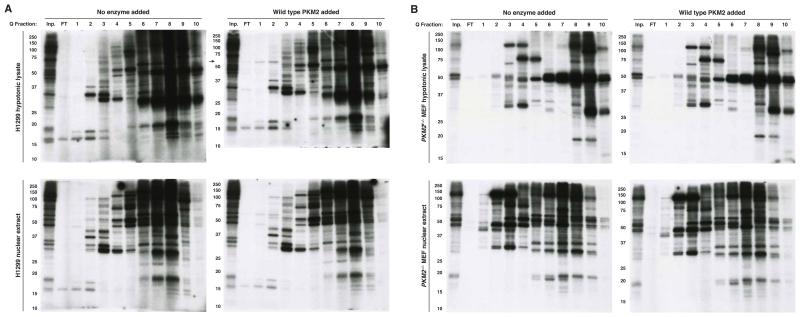
The role of pyruvate kinase M2 (PKM2) in cell proliferation is controversial. A unique function of PKM2 proposed to be important for the proliferation of some cancer cells involves the direct activity of this enzyme as a protein kinase; however, a detailed biochemical characterization of this activity is lacking. Using [(32)P]-phosphoenolpyruvate (PEP) we examine the direct substrates of PKM2 using recombinant enzyme and in vitro systems where PKM2 is genetically deleted. Labeling of some protein species from [(32)P]-PEP can be observed; however, most were dependent on the presence of ADP, and none were dependent on the presence of PKM2. In addition, we also failed to observe PKM2-dependent transfer of phosphate from ATP directly to protein. These findings argue against a role for PKM2 as a protein kinase.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Carreras J, Climent F, Bartrons R, Pons G. Effect of vanadate on the formation and stability of the phosphoenzyme forms of 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase and of phosphoglucomutase. Biochimica et biophysica acta. 1982;705:238–242. - PubMed
-
- Christofk HR, Vander Heiden MG, Wu N, Asara JM, Cantley LC. Pyruvate kinase M2 is a phosphotyrosine-binding protein. Nature. 2008;452:181–186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
