

Comparative genomics reveals multistep pathogenesis of E2A-PBX1 acute lymphoblastic leukemia
- PMID: 26301816
- PMCID: PMC4588292
- DOI: 10.1172/JCI81158
Comparative genomics reveals multistep pathogenesis of E2A-PBX1 acute lymphoblastic leukemia
Abstract
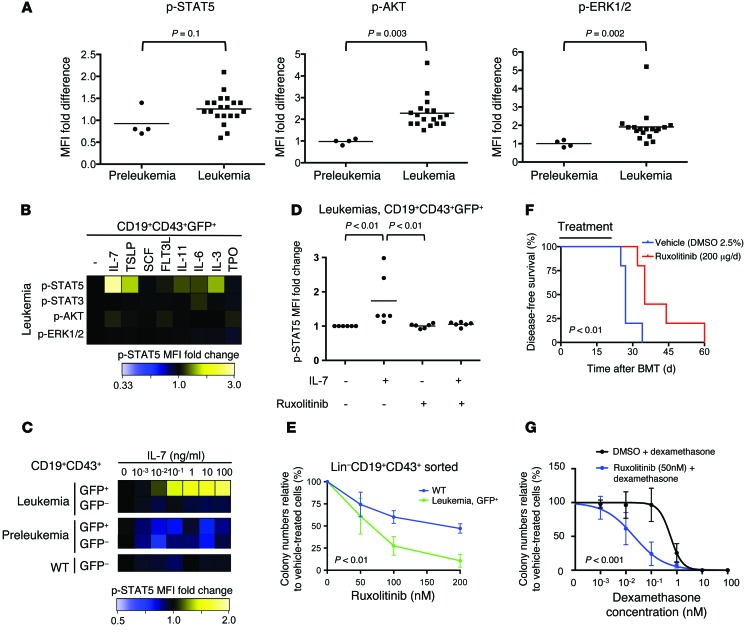
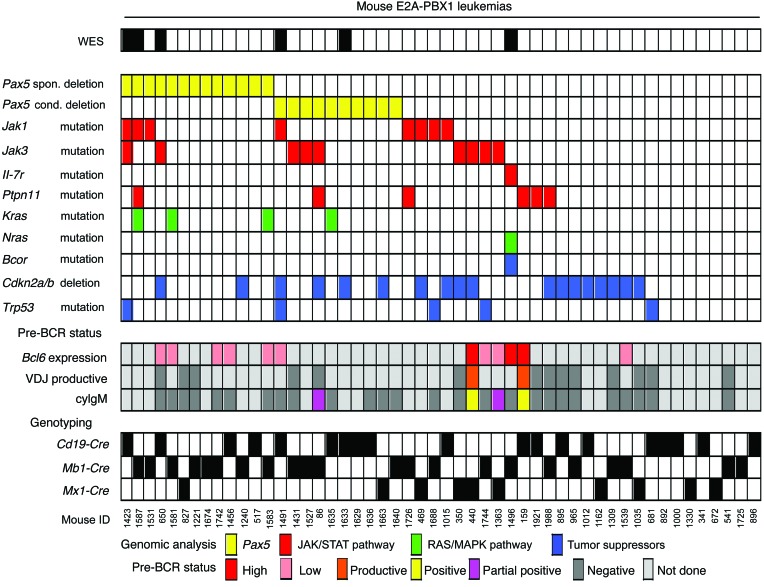
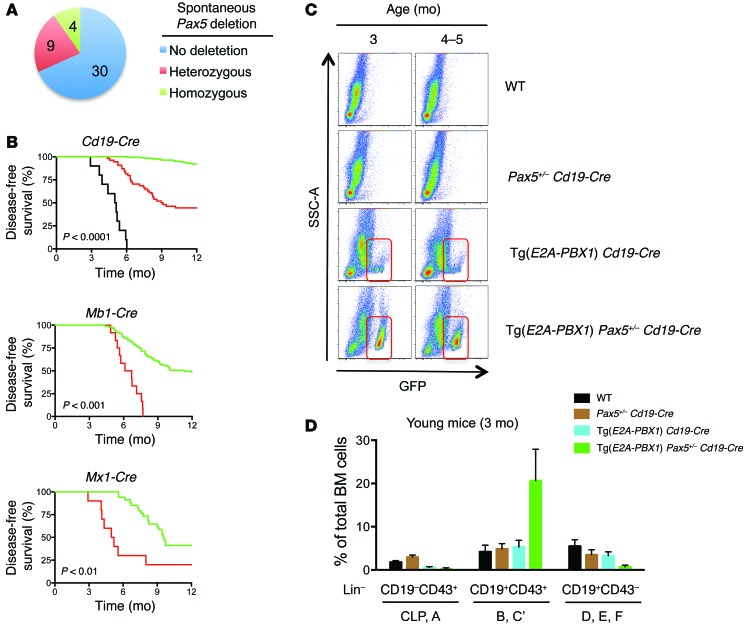
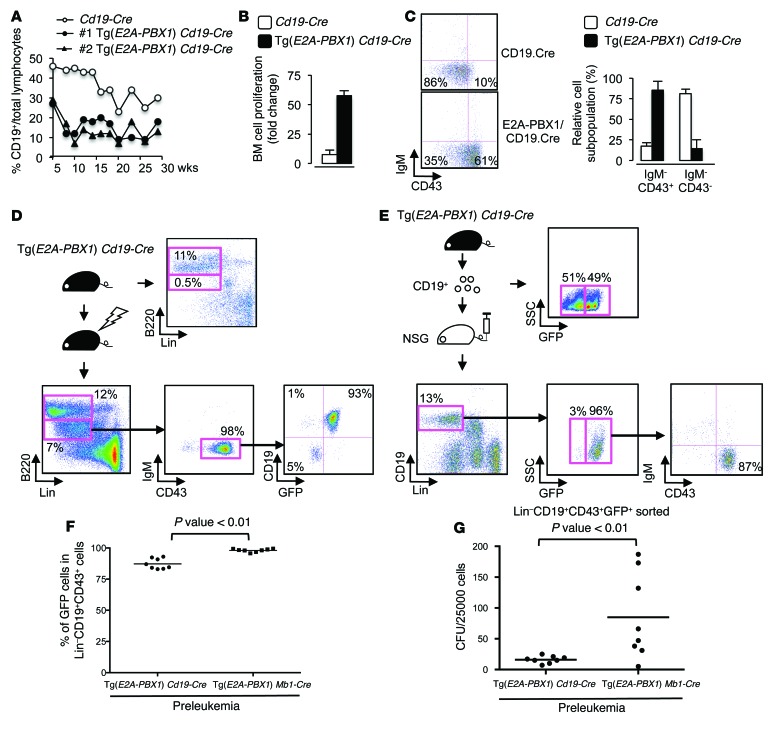
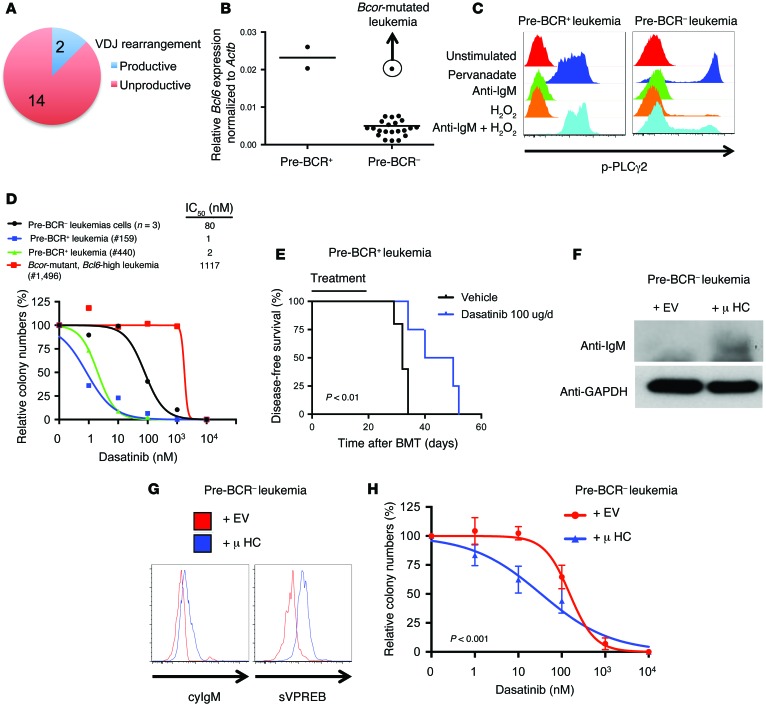
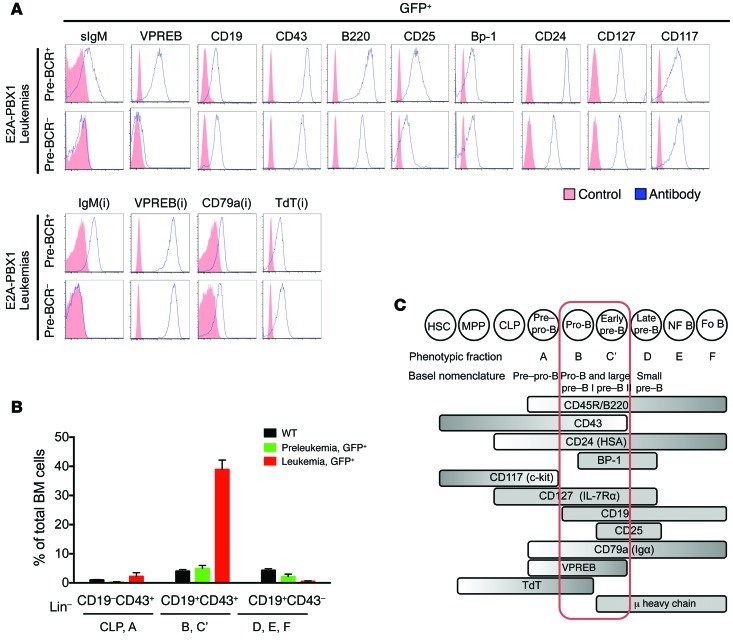
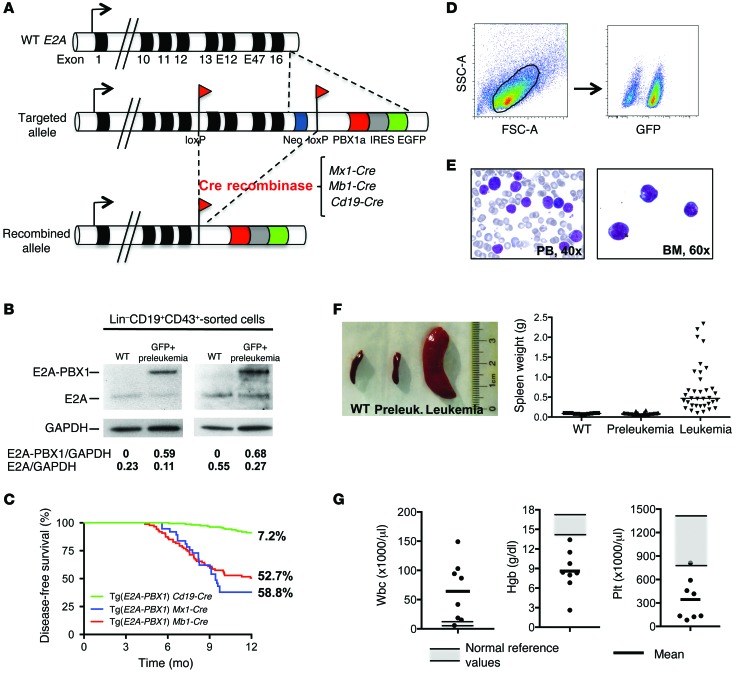
Acute lymphoblastic leukemia (ALL) is the most common childhood cancer; however, its genetic diversity limits investigation into the molecular pathogenesis of disease and development of therapeutic strategies. Here, we engineered mice that conditionally express the E2A-PBX1 fusion oncogene, which results from chromosomal translocation t(1;19) and is present in 5% to 7% of pediatric ALL cases. The incidence of leukemia in these mice varied from 5% to 50%, dependent on the Cre-driving promoter (Cd19, Mb1, or Mx1) used to induce E2A-PBX1 expression. Two distinct but highly similar subtypes of B cell precursor ALLs that differed by their pre-B cell receptor (pre-BCR) status were induced and displayed maturation arrest at the pro-B/large pre-B II stages of differentiation, similar to human E2A-PBX1 ALL. Somatic activation of E2A-PBX1 in B cell progenitors enhanced self-renewal and led to acquisition of multiple secondary genomic aberrations, including prominent spontaneous loss of Pax5. In preleukemic mice, conditional Pax5 deletion cooperated with E2A-PBX1 to expand progenitor B cell subpopulations, increasing penetrance and shortening leukemia latency. Recurrent secondary activating mutations were detected in key signaling pathways, most notably JAK/STAT, that leukemia cells require for proliferation. These data support conditional E2A-PBX1 mice as a model of human ALL and suggest targeting pre-BCR signaling and JAK kinases as potential therapeutic strategies.
Figures
Comment in
-
A robust in vivo model for B cell precursor acute lymphoblastic leukemia.J Clin Invest. 2015 Sep;125(9):3427-9. doi: 10.1172/JCI83799. Epub 2015 Aug 24. J Clin Invest. 2015. PMID: 26301807 Free PMC article. Review.
References
-
- Nowell P, Hungerford D. A minute chromosome in chronic granulocytic leukemia. Science. 1960;142:1497.
-
- Rowley JD. Editorial: The role of cytogenetics in hematology. Blood. 1976;48(1):1–7. - PubMed
-
- Mullighan CG. The molecular genetic makeup of acute lymphoblastic leukemia. Hematology Am Soc Hematol Educ Program. 2012;2012:389–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
