

Pyruvate carboxylation enables growth of SDH-deficient cells by supporting aspartate biosynthesis
- PMID: 26302408
- PMCID: PMC4591470
- DOI: 10.1038/ncb3233
Pyruvate carboxylation enables growth of SDH-deficient cells by supporting aspartate biosynthesis
Abstract
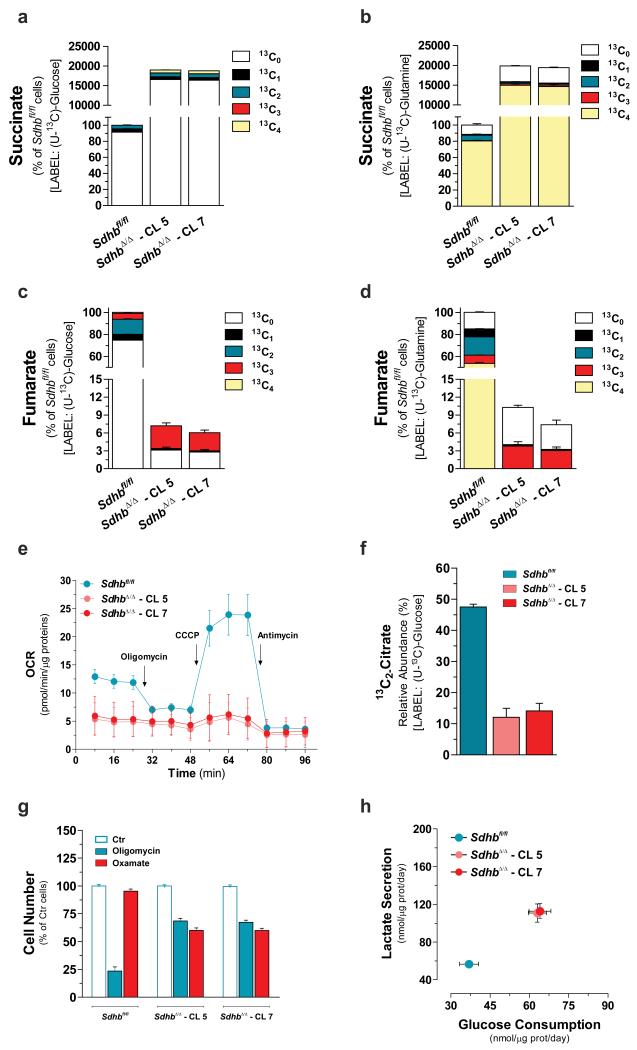
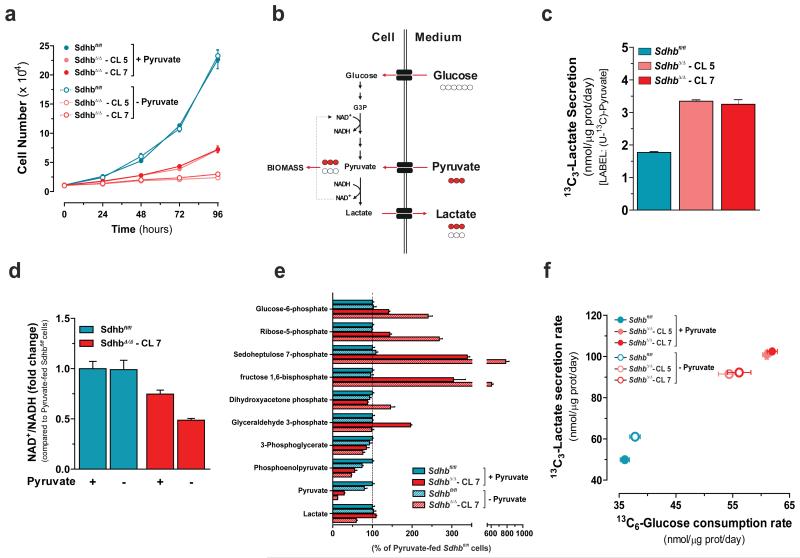
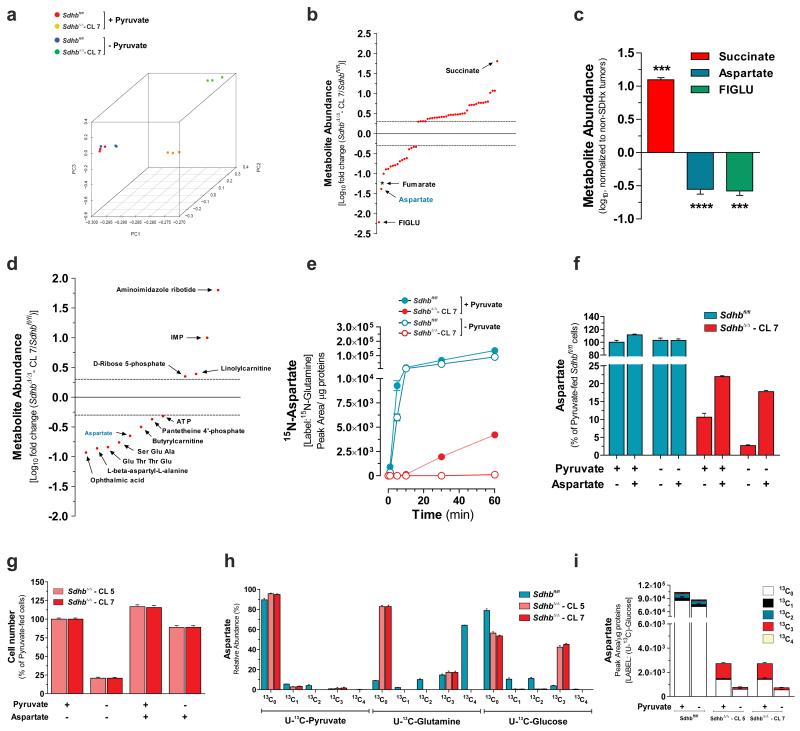
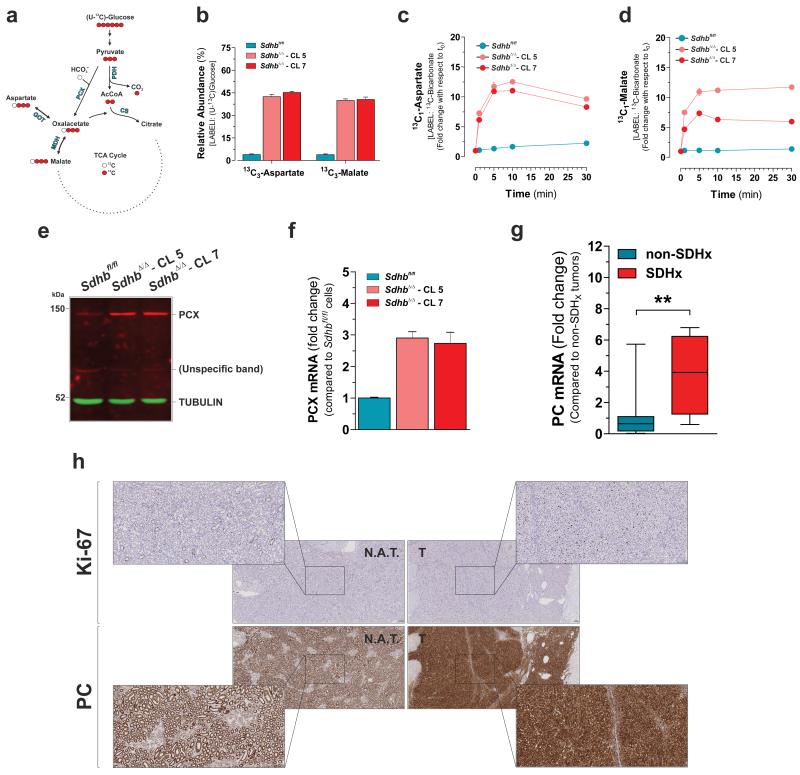
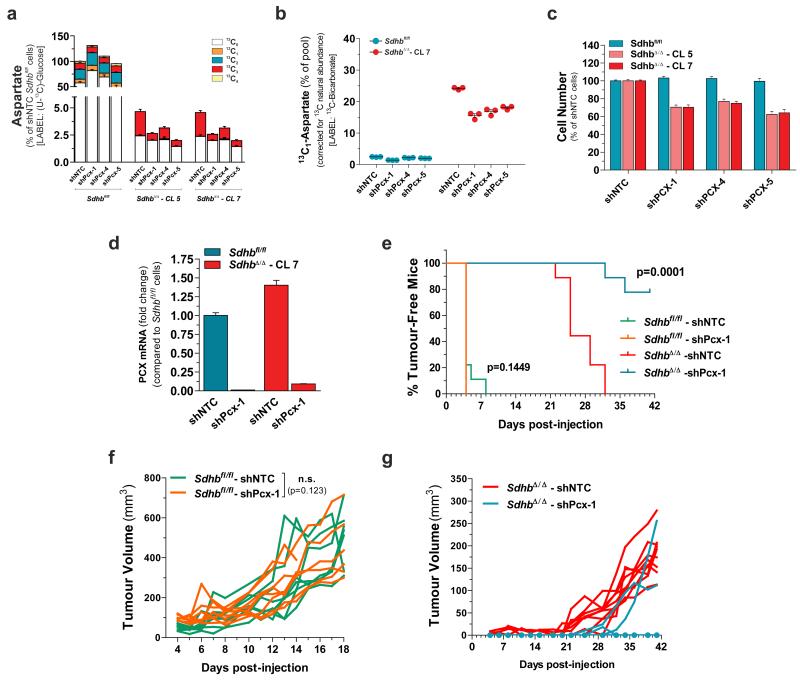
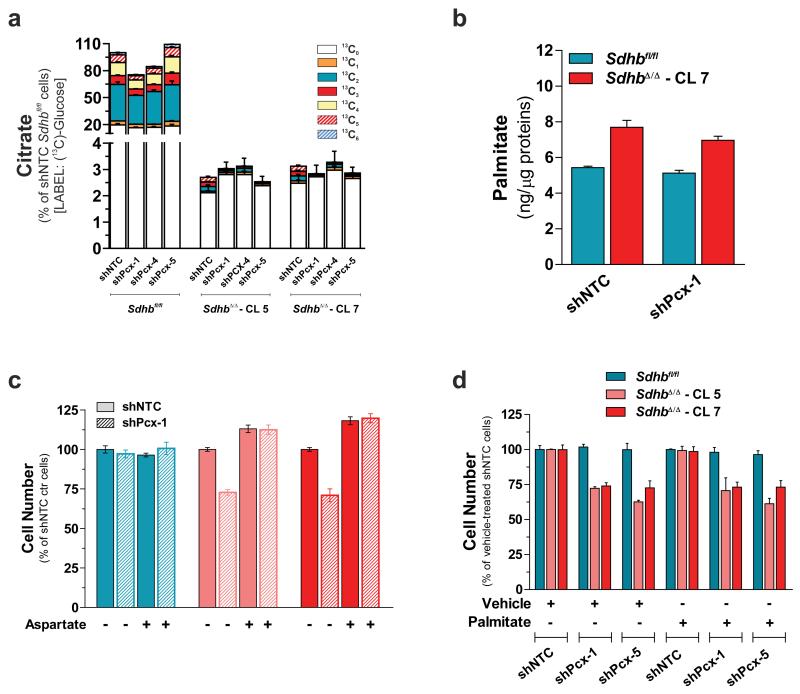
Succinate dehydrogenase (SDH) is a heterotetrameric nuclear-encoded complex responsible for the oxidation of succinate to fumarate in the tricarboxylic acid cycle. Loss-of-function mutations in any of the SDH genes are associated with cancer formation. However, the impact of SDH loss on cell metabolism and the mechanisms enabling growth of SDH-defective cells are largely unknown. Here, we generated Sdhb-ablated kidney mouse cells and used comparative metabolomics and stable-isotope-labelling approaches to identify nutritional requirements and metabolic adaptations to SDH loss. We found that lack of SDH activity commits cells to consume extracellular pyruvate, which sustains Warburg-like bioenergetic features. We further demonstrated that pyruvate carboxylation diverts glucose-derived carbons into aspartate biosynthesis, thus sustaining cell growth. By identifying pyruvate carboxylase as essential for the proliferation and tumorigenic capacity of SDH-deficient cells, this study revealed a metabolic vulnerability for potential future treatment of SDH-associated malignancies.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
