

Up-regulation of steroid biosynthesis by retinoid signaling: Implications for aging
- PMID: 26303142
- PMCID: PMC4583817
- DOI: 10.1016/j.mad.2015.08.007
Up-regulation of steroid biosynthesis by retinoid signaling: Implications for aging
Abstract
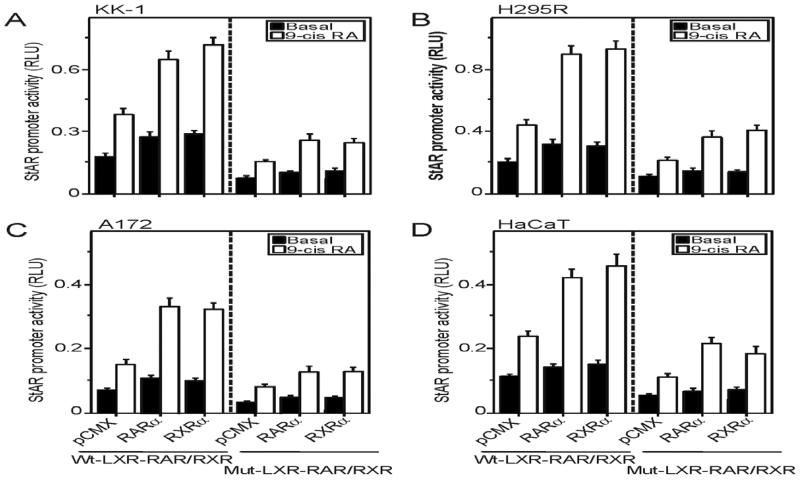
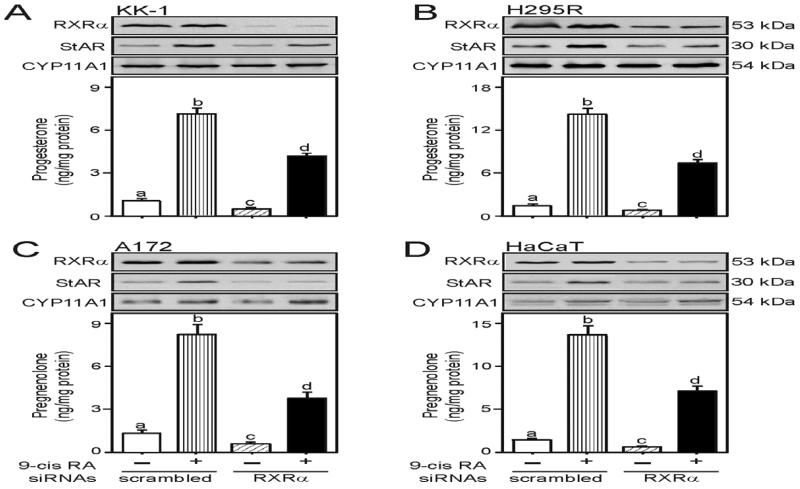
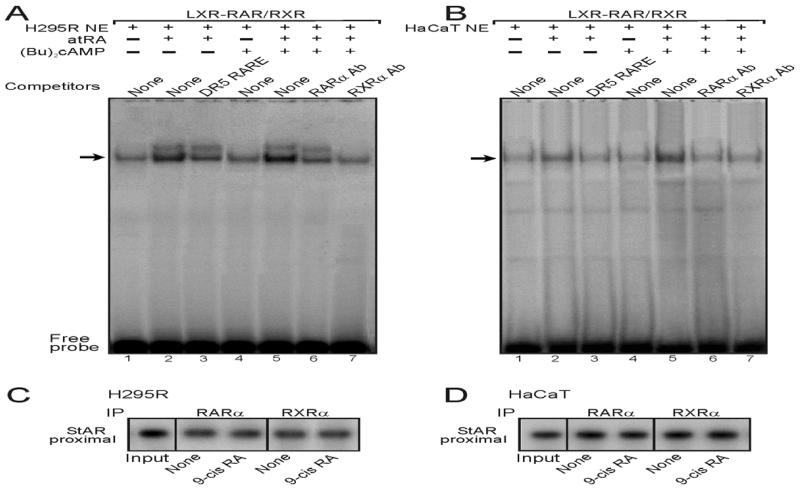
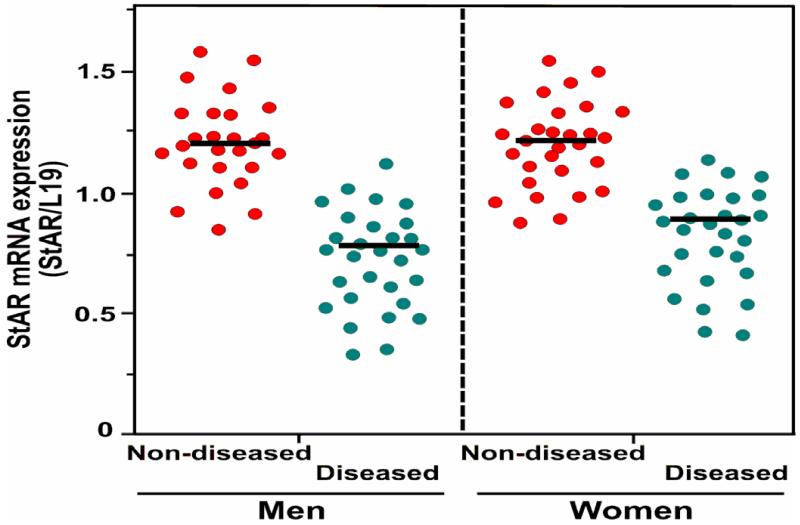
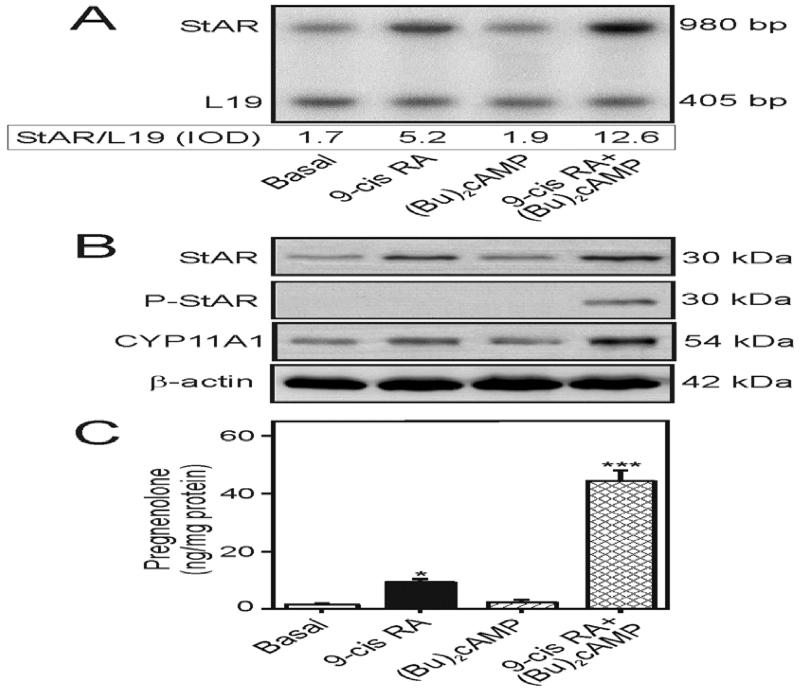
Retinoids (vitamin A and its derivatives) are critical for a spectrum of developmental and physiological processes, in which steroid hormones also play indispensable roles. The StAR protein predominantly regulates steroid biosynthesis in steroidogenic tissues. We have reported that regulation of retinoid, especially atRA and 9-cis RA, responsive StAR transcription is largely mediated by an LXR-RXR/RAR heterodimeric motif in the mouse StAR promoter. Herein we demonstrate that retinoids are capable of enhancing StAR protein, P-StAR, and steroid production in granulosa, adrenocortical, glial, and epidermal cells. Whereas transient expression of RARα and RXRα enhanced 9-cis RA induced StAR gene transcription, silencing of RXRα with siRNA, decreased StAR and steroid levels. An oligonucleotide probe encompassing an LXR-RXR/RAR motif bound to adrenocortical and epidermal keratinocyte nuclear proteins in EMSAs. ChIP studies revealed association of RARα and RXRα with the StAR proximal promoter. Further studies demonstrated that StAR mRNA levels decreased in diseased and elderly men and women skin tissues and that atRA could restore steroidogenesis in epidermal keratinocytes of aged individuals. These findings provide novel insights into the relevance of retinoid signaling in the up-regulation of steroid biosynthesis in various target tissues, and indicate that retinoid therapy may have important implications in age-related complications and diseases.
Keywords: RAR; RXR; Retinoids; StAR expression; Steroid biosynthesis; Steroidogenic cells; cAMP/PKA.
Copyright © 2015 Elsevier Ireland Ltd. All rights reserved.
Figures


References
-
- Bastien J, Adam-Stitah S, Plassat JL, Chambon P, Rochette-Egly C. The phosphorylation site located in the A region of retinoic X receptor alpha is required for the antiproliferative effect of retinoic acid (RA) and the activation of RA target genes in F9 cells. J. Biol. Chem. 2002;277:28683–28689. - PubMed
-
- Chahal HS, Drake WM. The endocrine system and ageing. J Pathol. 2007;211:173–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
