

Wiskott-Aldrich syndrome proteins in the nucleus: aWASH with possibilities
- PMID: 26305109
- PMCID: PMC4915506
- DOI: 10.1080/19491034.2015.1086051
Wiskott-Aldrich syndrome proteins in the nucleus: aWASH with possibilities
Abstract
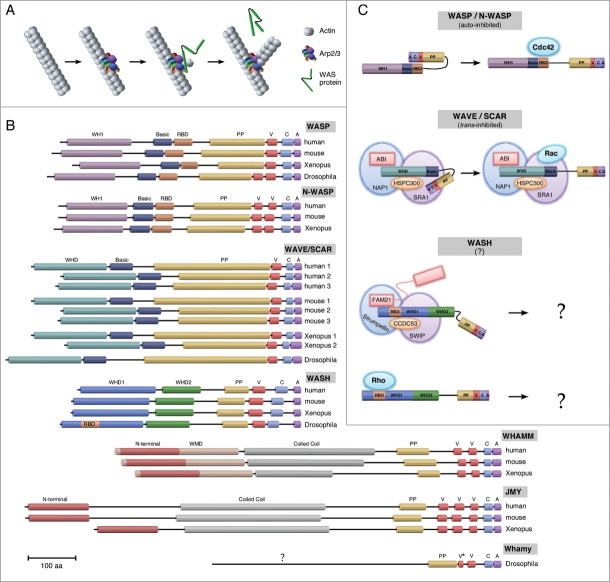
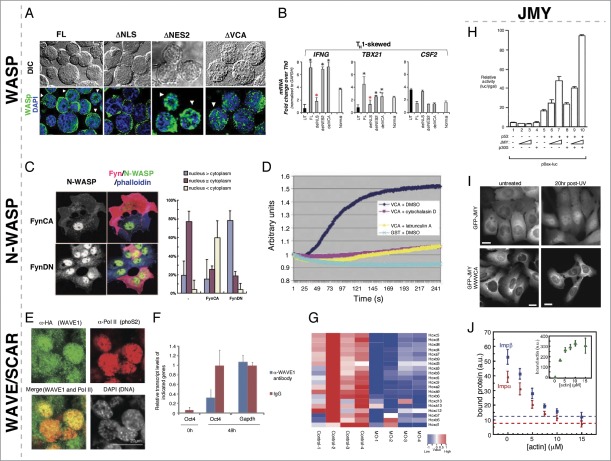
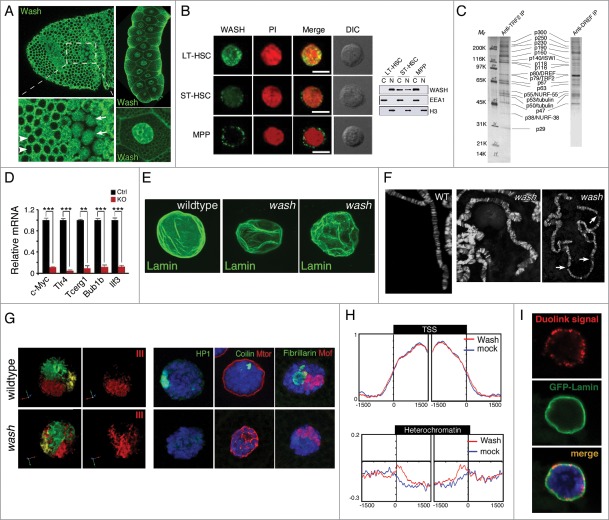
Actin and proteins that regulate its dynamics or interactions have well-established roles in the cytoplasm where they function as key components of the cytoskeleton to control diverse processes, including cellular infrastructure, cellular motility, cell signaling, and vesicle transport. Recent work has also uncovered roles for actin and its regulatory proteins in the nucleus, primarily in mechanisms governing gene expression. The Wiskott Aldrich Syndrome (WAS) family of proteins, comprising the WASP/N-WASP, SCAR/WAVE, WHAMM/JMY/WHAMY, and WASH subfamilies, function in the cytoplasm where they activate the Arp2/3 complex to form branched actin filaments. WAS proteins are present in the nucleus and have been implicated as transcriptional regulators. We found that Drosophila Wash, in addition to transcriptional effects, is involved in global nuclear architecture. Here we summarize the regulation and function of nuclear WAS proteins, and highlight how our work with Wash expands the possibilities for the functions of these proteins in the nucleus.
Keywords: JMY; WASH; WASP/N-WASP; WAVE/SCAR; WHAMM; Wiskott-Aldrich Syndrome; actin; nuclear architecture; nuclear organization; transcription.
Figures
Comment on
-
Wash interacts with lamin and affects global nuclear organization.Curr Biol. 2015 Mar 16;25(6):804-810. doi: 10.1016/j.cub.2015.01.052. Epub 2015 Mar 5. Curr Biol. 2015. PMID: 25754639 Free PMC article.
References
-
- Bezanilla M, Gladfelter AS, Kovar DR, Lee WL. Cytoskeletal dynamics: A view from the membrane. J Cell Biol 2015; 209:329-37; PMID:25963816; http://dx.doi.org/10.1083/jcb.201502062 - DOI - PMC - PubMed
-
- Burianek LE, Soderling SH. Under lock and key: spatiotemporal regulation of WASP family proteins coordinates separate dynamic cellular processes. Semin Cell Dev Biol 2013; 24:258-66; PMID:23291261; http://dx.doi.org/10.1016/j.semcdb.2012.12.005 - DOI - PMC - PubMed
-
- Takenawa T, Suetsugu S. The WASP-WAVE protein network: connecting the membrane to the cytoskeleton. Nat Rev Mol Cell Biol 2007; 8:37-48; PMID:17183359; http://dx.doi.org/10.1038/nrm2069 - DOI - PubMed
-
- Rottner K, Hanisch J, Campellone KG. WASH, WHAMM and JMY: regulation of Arp2/3 complex and beyond. Trends Cell Biol 2010; 20:650-61; PMID:20888769; http://dx.doi.org/10.1016/j.tcb.2010.08.014 - DOI - PubMed
-
- Veltman DM, Insall RH. WASP family proteins: their evolution and its physiological implications. Mol Biol Cell 2010; 21:2880-93; PMID:20573979; http://dx.doi.org/10.1091/mbc.E10-04-0372 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases