

Reduced Crossover Interference and Increased ZMM-Independent Recombination in the Absence of Tel1/ATM
- PMID: 26305689
- PMCID: PMC4549261
- DOI: 10.1371/journal.pgen.1005478
Reduced Crossover Interference and Increased ZMM-Independent Recombination in the Absence of Tel1/ATM
Erratum in
-
Correction: Reduced crossover interference and increased ZMM-independent recombination in the absence of Tel1/ATM.PLoS Genet. 2025 Jul 7;21(7):e1011779. doi: 10.1371/journal.pgen.1011779. eCollection 2025 Jul. PLoS Genet. 2025. PMID: 40622894 Free PMC article.
Abstract
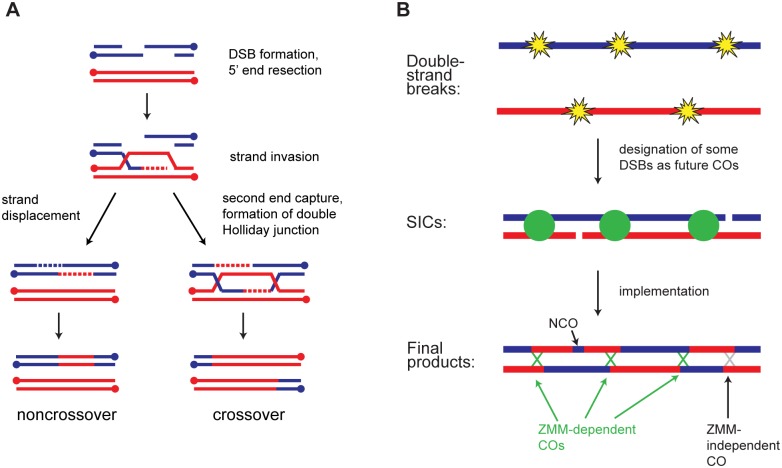
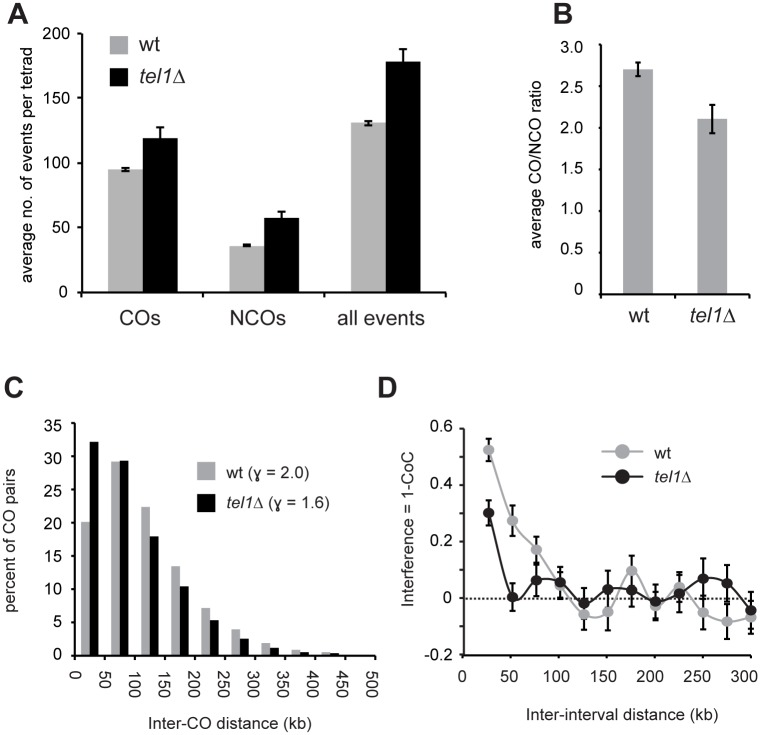
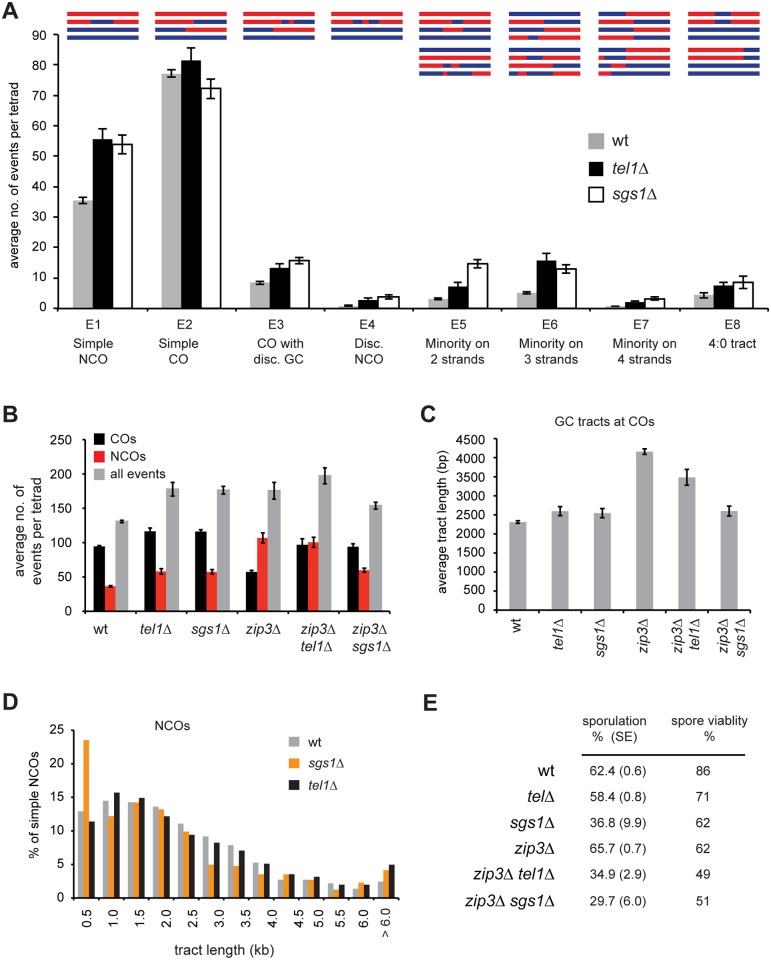
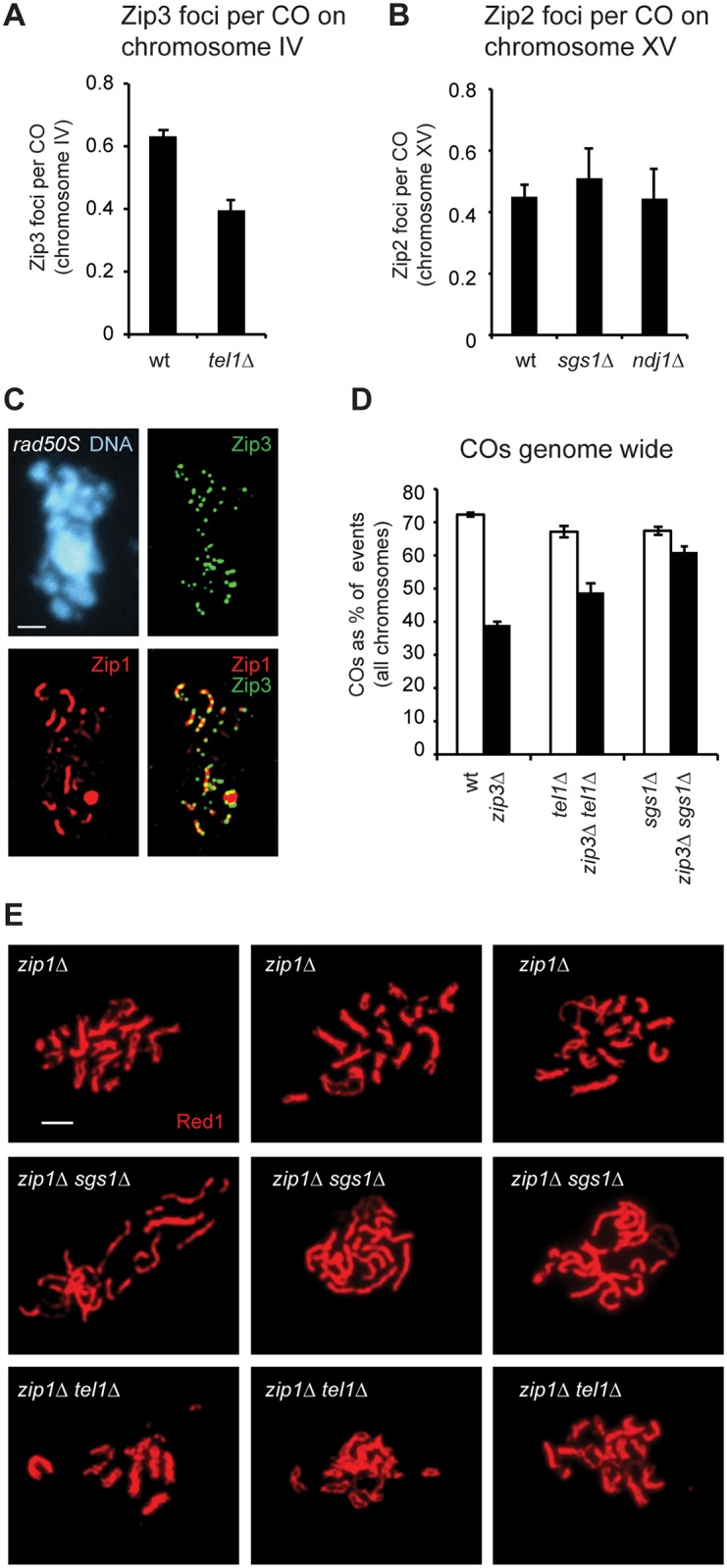
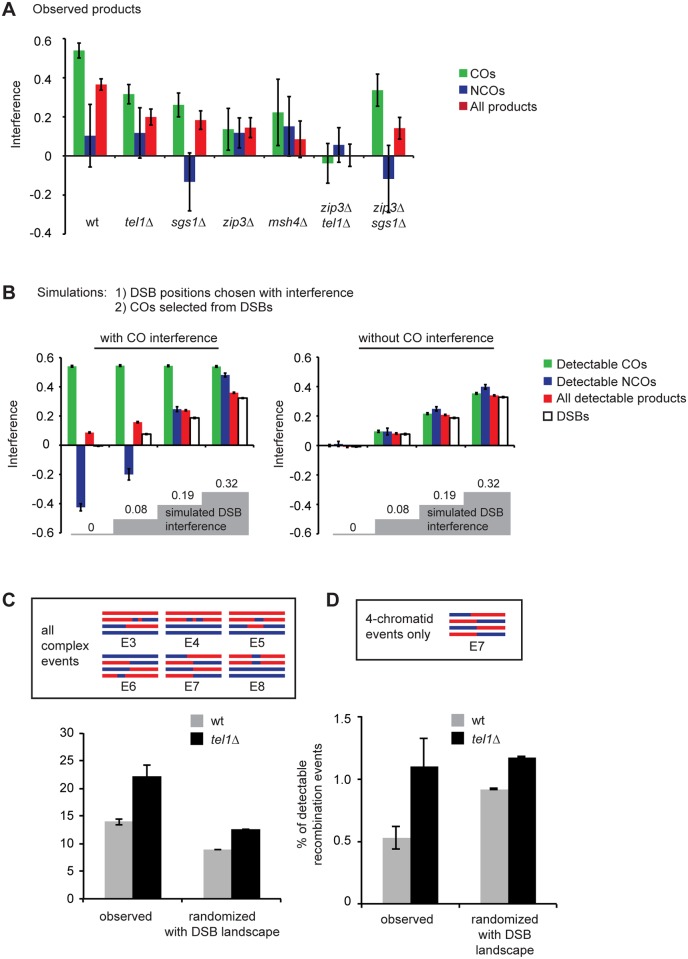
Meiotic recombination involves the repair of double-strand break (DSB) precursors as crossovers (COs) or noncrossovers (NCOs). The proper number and distribution of COs is critical for successful chromosome segregation and formation of viable gametes. In budding yeast the majority of COs occurs through a pathway dependent on the ZMM proteins (Zip2-Zip3-Zip4-Spo16, Msh4-Msh5, Mer3), which form foci at CO-committed sites. Here we show that the DNA-damage-response kinase Tel1/ATM limits ZMM-independent recombination. By whole-genome mapping of recombination products, we find that lack of Tel1 results in higher recombination and reduced CO interference. Yet the number of Zip3 foci in tel1Δ cells is similar to wild type, and these foci show normal interference. Analysis of recombination in a tel1Δ zip3Δ double mutant indicates that COs are less dependent on Zip3 in the absence of Tel1. Together these results reveal that in the absence of Tel1, a significant proportion of COs occurs through a non-ZMM-dependent pathway, contributing to a CO landscape with poor interference. We also see a significant change in the distribution of all detectable recombination products in the absence of Tel1, Sgs1, Zip3, or Msh4, providing evidence for altered DSB distribution. These results support the previous finding that DSB interference depends on Tel1, and further suggest an additional level of DSB interference created through local repression of DSBs around CO-designated sites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Sturtevant AH (1913) A Third Group of Linked Genes in Drosophila Ampelophila . Science 37: 990–992. - PubMed
-
- Allers T, Lichten M (2001) Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106: 47–57. - PubMed
-
- Borner GV, Kleckner N, Hunter N (2004) Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell 117: 29–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
