

The Cell Shape-determining Csd6 Protein from Helicobacter pylori Constitutes a New Family of L,D-Carboxypeptidase
- PMID: 26306031
- PMCID: PMC4599014
- DOI: 10.1074/jbc.M115.658781
The Cell Shape-determining Csd6 Protein from Helicobacter pylori Constitutes a New Family of L,D-Carboxypeptidase
Abstract
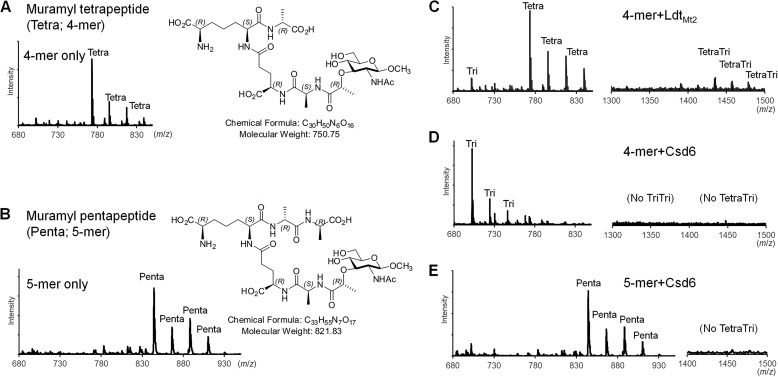
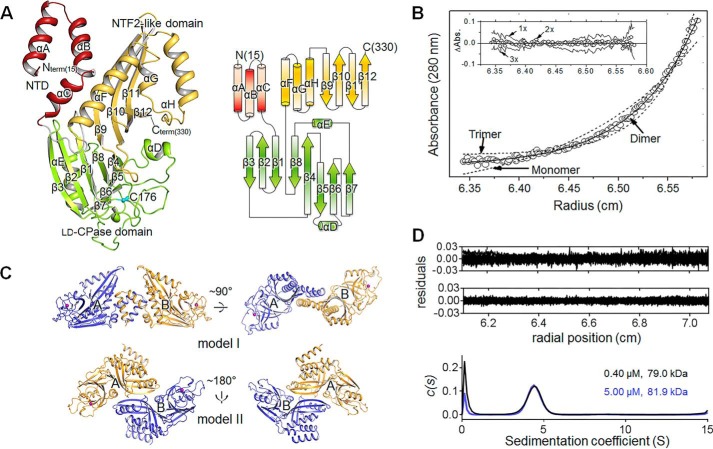
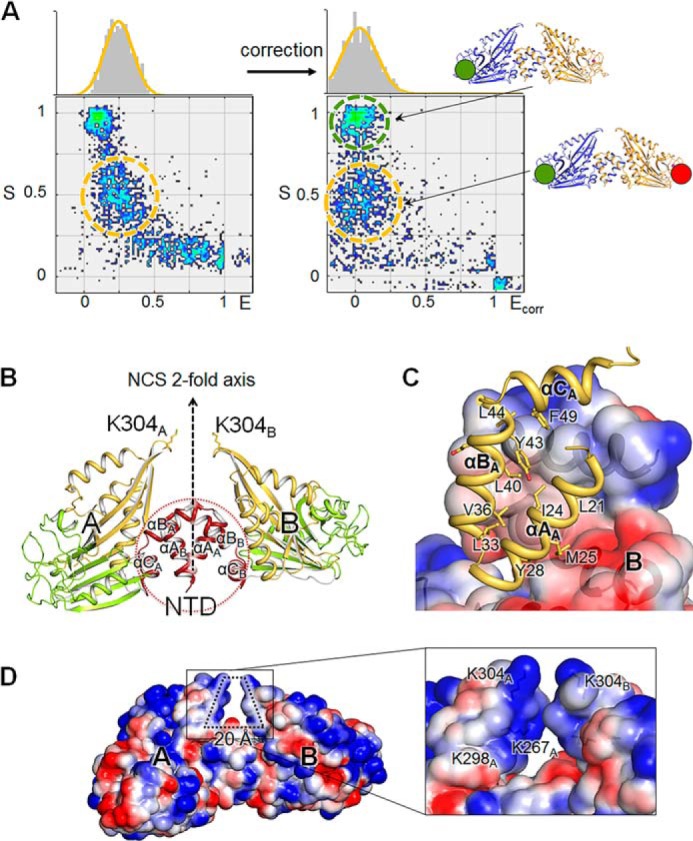
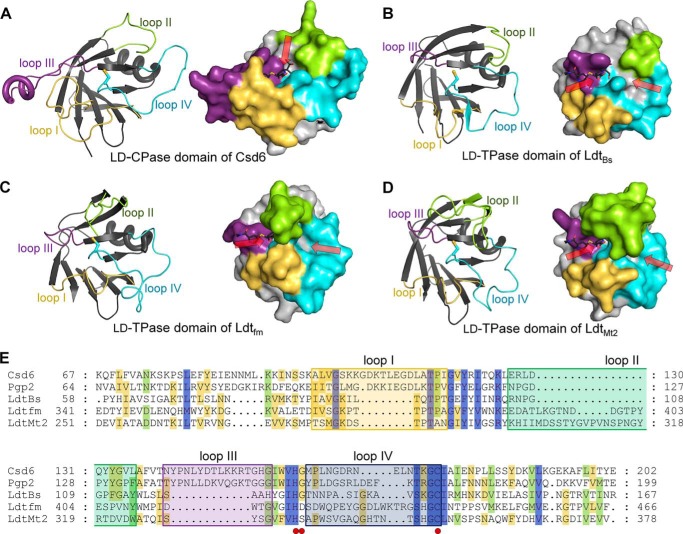
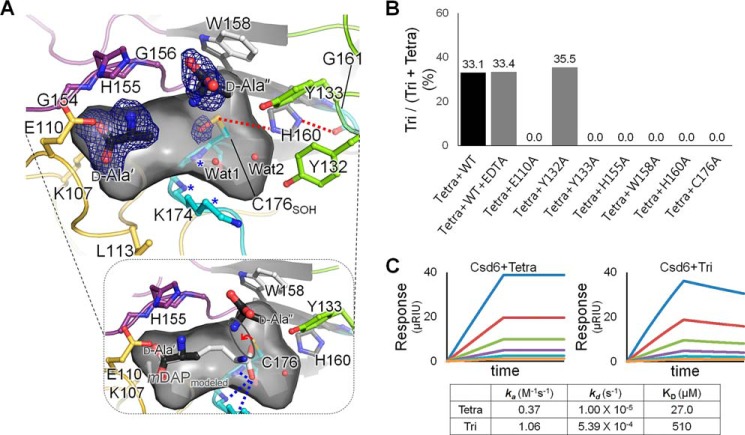
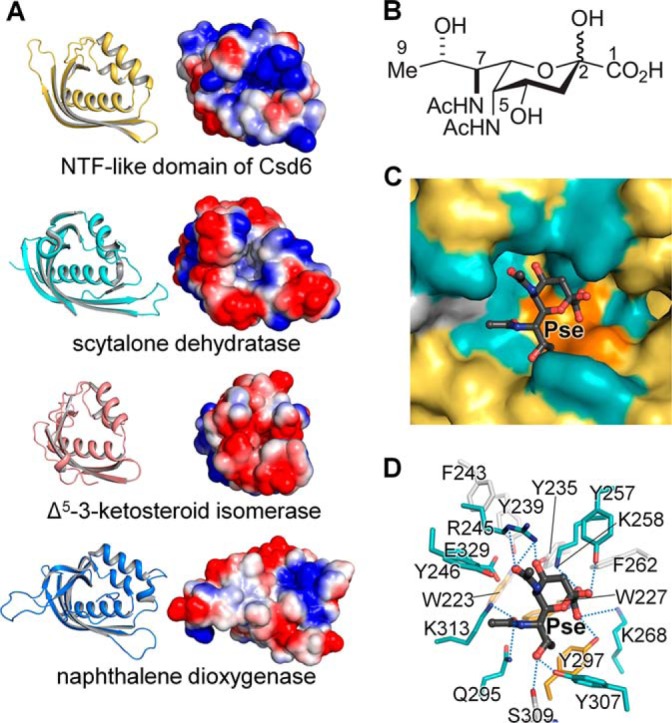
Helicobacter pylori causes gastrointestinal diseases, including gastric cancer. Its high motility in the viscous gastric mucosa facilitates colonization of the human stomach and depends on the helical cell shape and the flagella. In H. pylori, Csd6 is one of the cell shape-determining proteins that play key roles in alteration of cross-linking or by trimming of peptidoglycan muropeptides. Csd6 is also involved in deglycosylation of the flagellar protein FlaA. To better understand its function, biochemical, biophysical, and structural characterizations were carried out. We show that Csd6 has a three-domain architecture and exists as a dimer in solution. The N-terminal domain plays a key role in dimerization. The middle catalytic domain resembles those of l,d-transpeptidases, but its pocket-shaped active site is uniquely defined by the four loops I to IV, among which loops I and III show the most distinct variations from the known l,d-transpeptidases. Mass analyses confirm that Csd6 functions only as an l,d-carboxypeptidase and not as an l,d-transpeptidase. The d-Ala-complexed structure suggests possible binding modes of both the substrate and product to the catalytic domain. The C-terminal nuclear transport factor 2-like domain possesses a deep pocket for possible binding of pseudaminic acid, and in silico docking supports its role in deglycosylation of flagellin. On the basis of these findings, it is proposed that H. pylori Csd6 and its homologs constitute a new family of l,d-carboxypeptidase. This work provides insights into the function of Csd6 in regulating the helical cell shape and motility of H. pylori.
Keywords: Csd6; HP0518; Helicobacter pylori; L,D-carboxypeptidase; cell motility; cell shape; flagellin; peptidoglycan; protein structure; structure-function.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Structural basis for the recognition of muramyltripeptide by Helicobacter pylori Csd4, a D,L-carboxypeptidase controlling the helical cell shape.Acta Crystallogr D Biol Crystallogr. 2014 Nov;70(Pt 11):2800-12. doi: 10.1107/S1399004714018732. Epub 2014 Oct 16. Acta Crystallogr D Biol Crystallogr. 2014. PMID: 25372672 Free PMC article.
-
Structural Basis of the Heterodimer Formation between Cell Shape-Determining Proteins Csd1 and Csd2 from Helicobacter pylori.PLoS One. 2016 Oct 6;11(10):e0164243. doi: 10.1371/journal.pone.0164243. eCollection 2016. PLoS One. 2016. PMID: 27711177 Free PMC article.
-
Structure of Csd3 from Helicobacter pylori, a cell shape-determining metallopeptidase.Acta Crystallogr D Biol Crystallogr. 2015 Mar;71(Pt 3):675-86. doi: 10.1107/S1399004715000152. Epub 2015 Feb 26. Acta Crystallogr D Biol Crystallogr. 2015. PMID: 25760614 Free PMC article.
-
Role of Flagella in the Pathogenesis of Helicobacter pylori.Curr Microbiol. 2017 Jul;74(7):863-869. doi: 10.1007/s00284-017-1256-4. Epub 2017 Apr 26. Curr Microbiol. 2017. PMID: 28444418 Free PMC article. Review.
-
Motility of Different Gastric Helicobacter spp.Microorganisms. 2023 Mar 1;11(3):634. doi: 10.3390/microorganisms11030634. Microorganisms. 2023. PMID: 36985208 Free PMC article. Review.
Cited by
-
Nonhelical Helicobacter pylori Mutants Show Altered Gland Colonization and Elicit Less Gastric Pathology than Helical Bacteria during Chronic Infection.Infect Immun. 2019 Jun 20;87(7):e00904-18. doi: 10.1128/IAI.00904-18. Print 2019 Jul. Infect Immun. 2019. PMID: 31061142 Free PMC article.
-
Isolation, Purification and Characterization of L,D-transpeptidase 2 from Mycobacterium tuberculosis.Acta Naturae. 2019 Jan-Mar;11(1):23-28. Acta Naturae. 2019. PMID: 31024745 Free PMC article.
-
Morphological change of coiled bacterium Spirosoma linguale with acquisition of β-lactam resistance.Sci Rep. 2021 Jun 24;11(1):13278. doi: 10.1038/s41598-021-92787-8. Sci Rep. 2021. PMID: 34168257 Free PMC article.
-
Cell Wall Hydrolases in Bacteria: Insight on the Diversity of Cell Wall Amidases, Glycosidases and Peptidases Toward Peptidoglycan.Front Microbiol. 2019 Feb 28;10:331. doi: 10.3389/fmicb.2019.00331. eCollection 2019. Front Microbiol. 2019. PMID: 30873139 Free PMC article. Review.
-
Cleavage of Braun's lipoprotein Lpp from the bacterial peptidoglycan by a paralog of l,d-transpeptidases, LdtF.Proc Natl Acad Sci U S A. 2021 May 11;118(19):e2101989118. doi: 10.1073/pnas.2101989118. Proc Natl Acad Sci U S A. 2021. PMID: 33941679 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
