

Chamber identity programs drive early functional partitioning of the heart
- PMID: 26306682
- PMCID: PMC4560818
- DOI: 10.1038/ncomms9146
Chamber identity programs drive early functional partitioning of the heart
Abstract
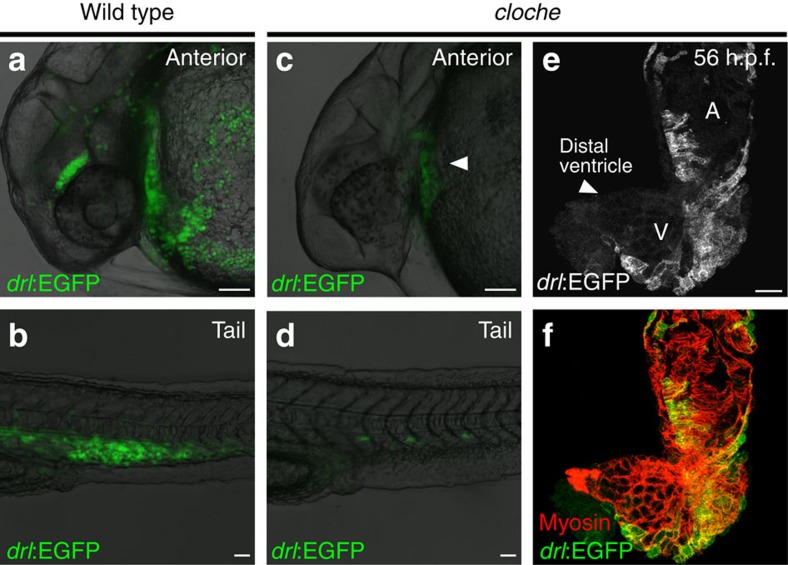
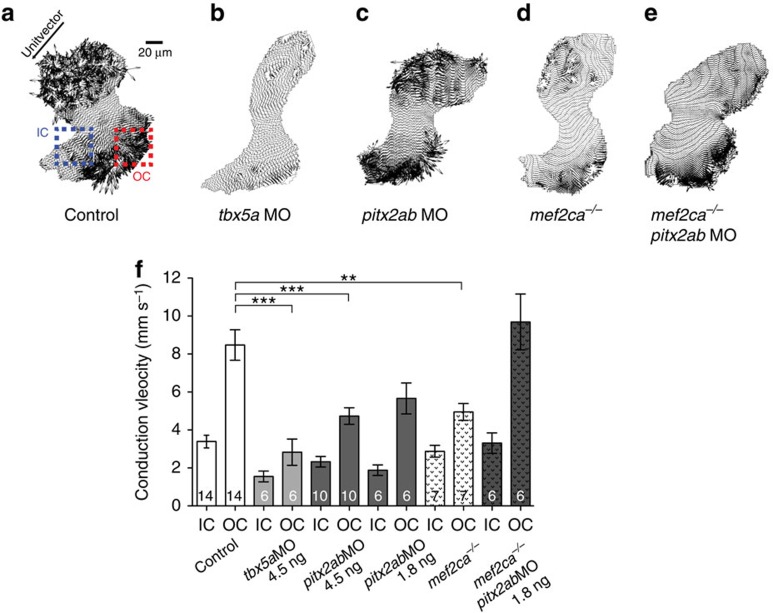
The vertebrate heart muscle (myocardium) develops from the first heart field (FHF) and expands by adding second heart field (SHF) cells. While both lineages exist already in teleosts, the primordial contributions of FHF and SHF to heart structure and function remain incompletely understood. Here we delineate the functional contribution of the FHF and SHF to the zebrafish heart using the cis-regulatory elements of the draculin (drl) gene. The drl reporters initially delineate the lateral plate mesoderm, including heart progenitors. Subsequent myocardial drl reporter expression restricts to FHF descendants. We harnessed this unique feature to uncover that loss of tbx5a and pitx2 affect relative FHF versus SHF contributions to the heart. High-resolution physiology reveals distinctive electrical properties of each heart field territory that define a functional boundary within the single zebrafish ventricle. Our data establish that the transcriptional program driving cardiac septation regulates physiologic ventricle partitioning, which successively provides mechanical advantages of sequential contraction.
Conflict of interest statement
L.I.Z. is a founder and stock holder of Fate, and a scientific advisor for Stemgent. The remaining authors declare no competing financial interests.
Figures




References
-
- Garrity D. M., Childs S. & Fishman M. C. The heartstrings mutation in zebrafish causes heart/fin Tbx5 deficiency syndrome. Development 129, 4635–4645 (2002). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
