

Suppression of different classes of somatic mutations in Arabidopsis by vir gene-expressing Agrobacterium strains
- PMID: 26307100
- PMCID: PMC4549908
- DOI: 10.1186/s12870-015-0595-1
Suppression of different classes of somatic mutations in Arabidopsis by vir gene-expressing Agrobacterium strains
Abstract
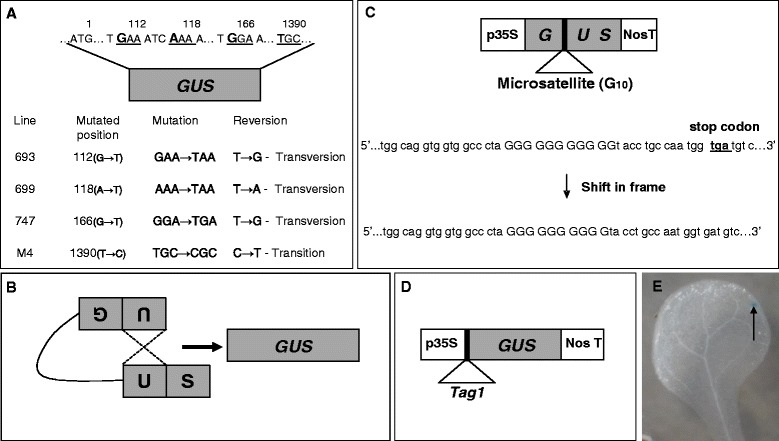
Background: Agrobacterium infection, which is widely used to generate transgenic plants, is often accompanied by T-DNA-linked mutations and transpositions in flowering plants. It is not known if Agrobacterium infection also affects the rates of point mutations, somatic homologous recombinations (SHR) and frame-shift mutations (FSM). We examined the effects of Agrobacterium infection on five types of somatic mutations using a set of mutation detector lines of Arabidopsis thaliana. To verify the effect of secreted factors, we exposed the plants to different Agrobacterium strains, including wild type (Ach5), its derivatives lacking vir genes, oncogenes or T-DNA, and the heat-killed form for 48 h post-infection; also, for a smaller set of strains, we examined the rates of three types of mutations at multiple time-points. The mutation detector lines carried a non-functional β-glucuronidase gene (GUS) and a reversion of mutated GUS to its functional form resulted in blue spots. Based on the number of blue spots visible in plants grown for a further two weeks, we estimated the mutation frequencies.
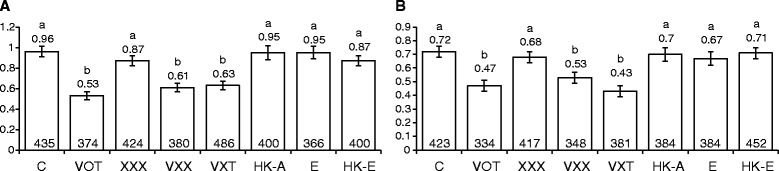
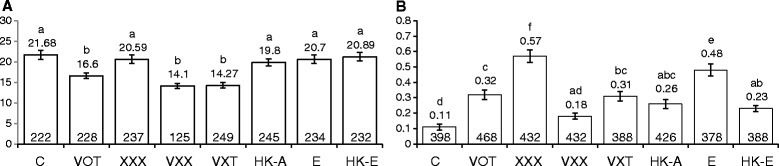
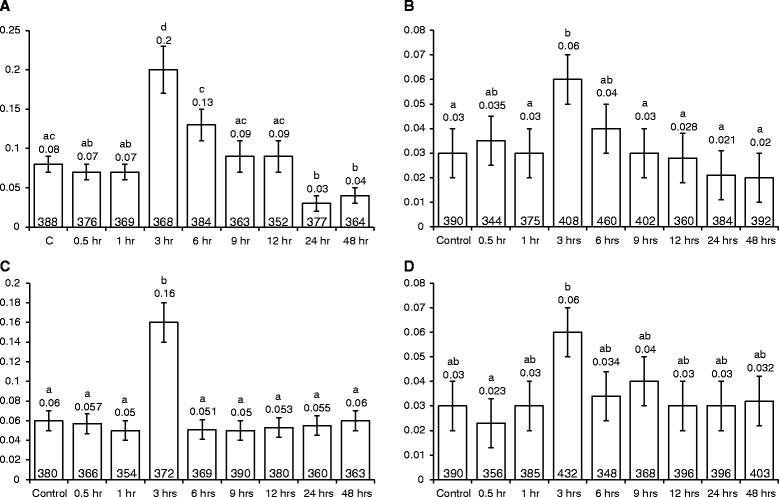
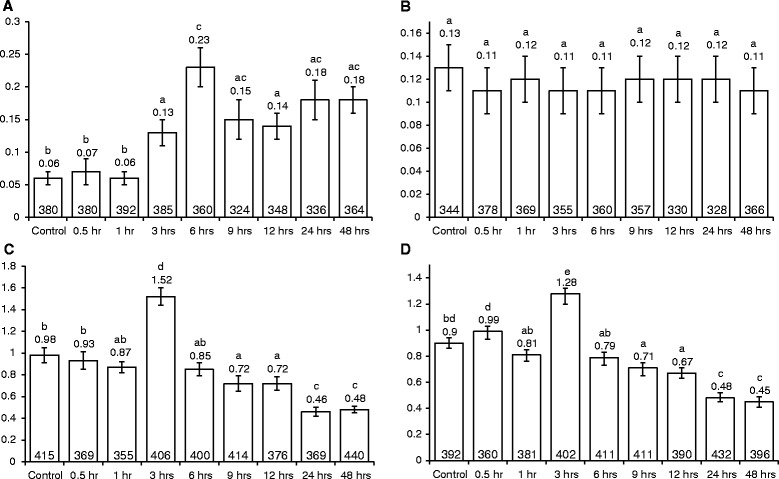
Results: For plants co-cultivated for 48 h with Agrobacterium, if the strain contained vir genes, then the rates of transversions, SHRs and FSMs (measured 2 weeks later) were lower than those of uninfected controls. In contrast, co-cultivation for 48 h with any of the Agrobacterium strains raised the transposition rates above control levels. The multiple time-point study showed that in seedlings co-cultivated with wild type Ach5, the reduced rates of transversions and SHRs after 48 h co-cultivation represent an apparent suppression of an earlier short-lived increase in mutation rates (peaking for plants co-cultivated for 3 h). An increase after 3 h co-cultivation was also seen for rates of transversions (but not SHR) in seedlings exposed to the strain lacking vir genes, oncogenes and T-DNA. However, the mutation rates in plants co-cultivated for longer times with this strain subsequently dropped below levels seen in uninfected controls, consistent with the results of the single time-point study.
Conclusions: The rates of various classes of mutations that result from Agrobacterium infection depend upon the duration of infection and the type of pathogen derived factors (such as Vir proteins, oncoproteins or T-DNA) possessed by the strain. Strains with vir genes, including the type used for plant transformation, suppressed selected classes of somatic mutations. Our study also provides evidence of a pathogen that can at least partly counter the induction of mutations in an infected plant.
Figures

Similar articles
-
pSa causes oncogenic suppression of Agrobacterium by inhibiting VirE2 protein export.J Bacteriol. 1999 Jan;181(1):186-96. doi: 10.1128/JB.181.1.186-196.1999. J Bacteriol. 1999. PMID: 9864329 Free PMC article.
-
Additional virulence genes and sonication enhance Agrobacterium tumefaciens-mediated loblolly pine transformation.Plant Cell Rep. 2003 Feb;21(6):555-62. doi: 10.1007/s00299-002-0550-0. Epub 2002 Nov 26. Plant Cell Rep. 2003. PMID: 12789430
-
Ethylene production in plants during transformation suppresses vir gene expression in Agrobacterium tumefaciens.New Phytol. 2008;178(3):647-56. doi: 10.1111/j.1469-8137.2008.02400.x. Epub 2008 Mar 6. New Phytol. 2008. PMID: 18331427
-
Arabidopsis thaliana floral dip transformation method.Methods Mol Biol. 2006;343:87-103. doi: 10.1385/1-59745-130-4:87. Methods Mol Biol. 2006. PMID: 16988336 Review.
-
Agrobacterium transformation of Arabidopsis thaliana roots: a quantitative assay.Methods Mol Biol. 2006;343:105-13. doi: 10.1385/1-59745-130-4:105. Methods Mol Biol. 2006. PMID: 16988337 Review.
Cited by
-
Screening internal controls for expression analyses involving numerous treatments by combining statistical methods with reference gene selection tools.Physiol Mol Biol Plants. 2019 Jan;25(1):289-301. doi: 10.1007/s12298-018-0608-2. Epub 2018 Oct 9. Physiol Mol Biol Plants. 2019. PMID: 30804650 Free PMC article.
-
Essential role of the BRCA2B gene in somatic homologous recombination in Arabidopsis thaliana.BioTechnologia (Pozn). 2023 Dec 21;104(4):371-380. doi: 10.5114/bta.2023.132773. eCollection 2023. BioTechnologia (Pozn). 2023. PMID: 38213474 Free PMC article.
References
-
- Escobar MA, Dandekar AM. Agrobacterium tumefaciens as an agent of disease. Trends Plant Sci. 2003;8:380–6. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
