

Tomato R2R3-MYB Proteins SlANT1 and SlAN2: Same Protein Activity, Different Roles
- PMID: 26308527
- PMCID: PMC4556288
- DOI: 10.1371/journal.pone.0136365
Tomato R2R3-MYB Proteins SlANT1 and SlAN2: Same Protein Activity, Different Roles
Abstract
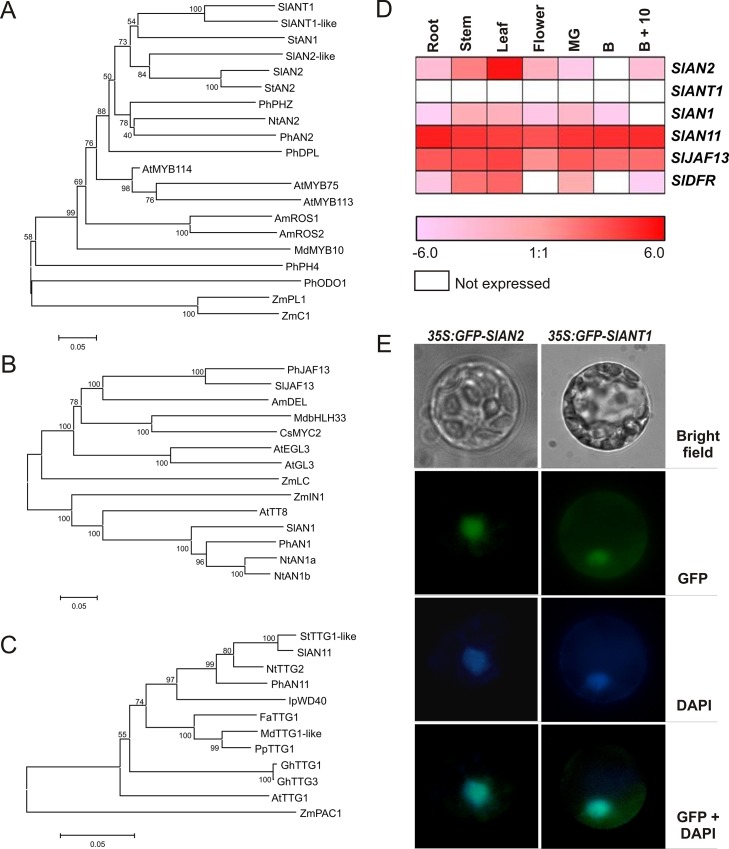
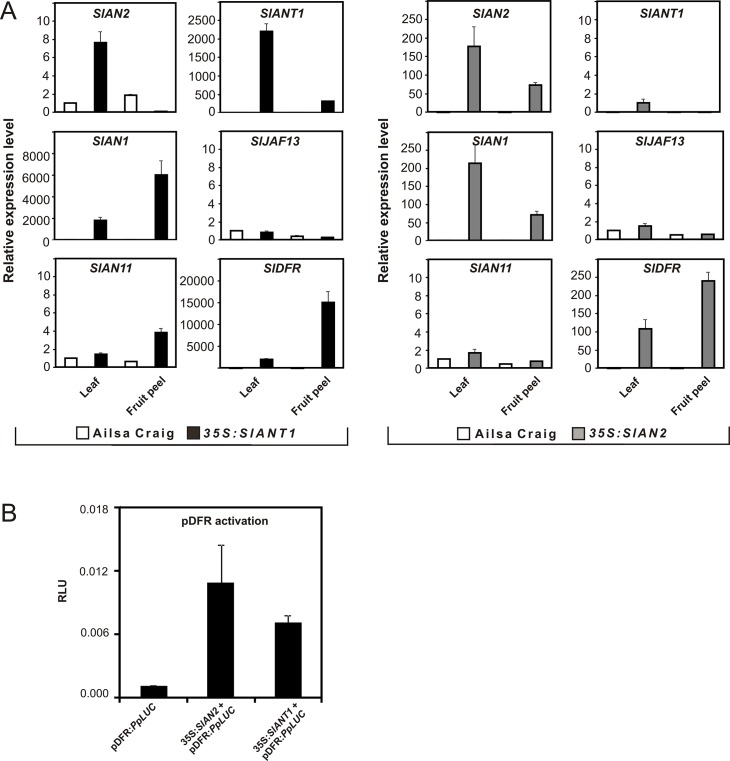
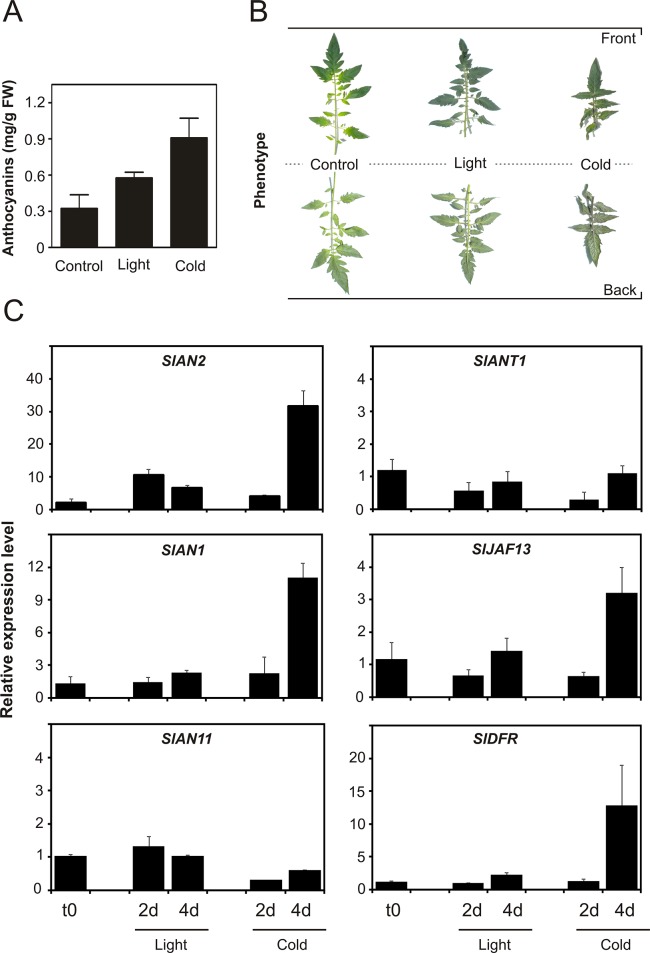
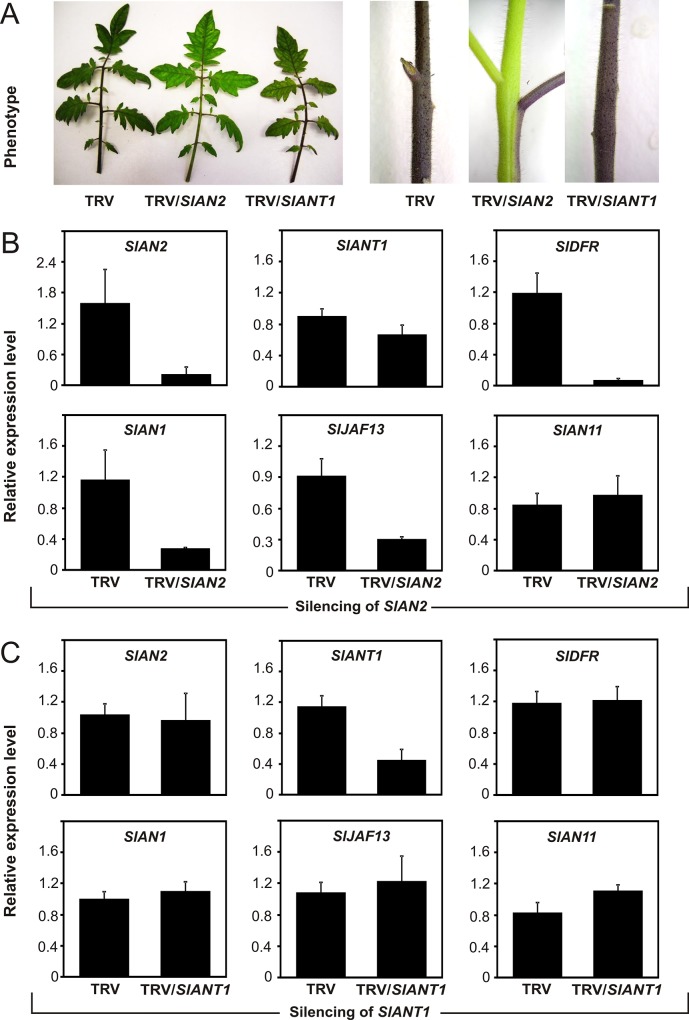
Anthocyanins are water-soluble polyphenolic compounds with a high nutraceutical value. Despite the fact that cultivated tomato varieties do not accumulate anthocyanins in the fruit, the biosynthetic pathway can be activated in the vegetative organs by several environmental stimuli. Little is known about the molecular mechanisms regulating anthocyanin synthesis in tomato. Here, we carried out a molecular and functional characterization of two genes, SlAN2 and SlANT1, encoding two R2R3-MYB transcription factors. We show that both can induce ectopic anthocyanin synthesis in transgenic tomato lines, including the fruit. However, only SlAN2 acts as a positive regulator of anthocyanin synthesis in vegetative tissues under high light or low temperature conditions.
Conflict of interest statement
Figures

References
-
- Koes R, Verweij CW, Quattrocchio F. Flavonoids: a colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005; 5: 236–242. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
