

The C9orf72 repeat expansion disrupts nucleocytoplasmic transport
- PMID: 26308891
- PMCID: PMC4800742
- DOI: 10.1038/nature14973
The C9orf72 repeat expansion disrupts nucleocytoplasmic transport
Abstract
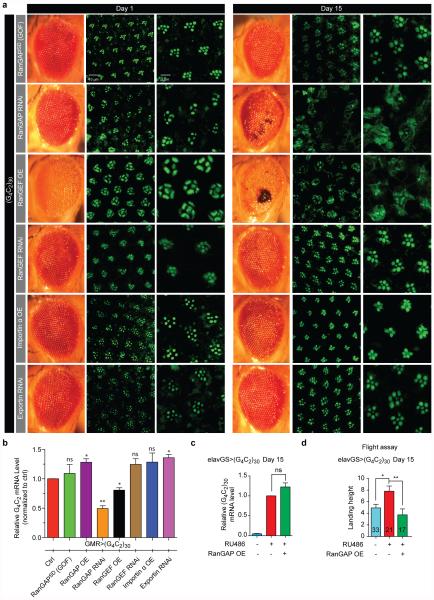
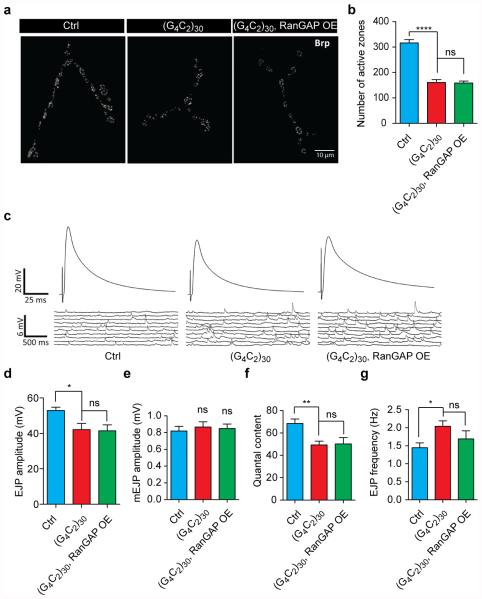
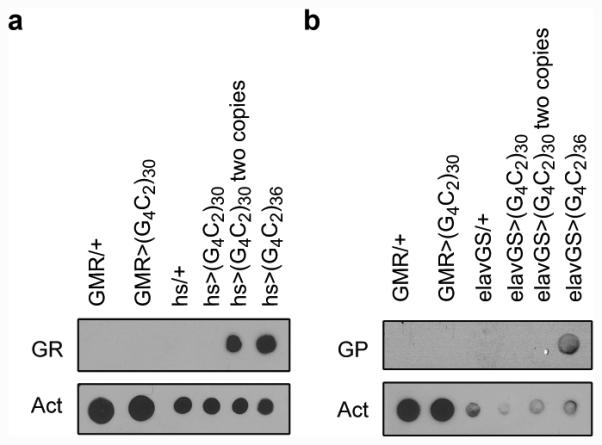
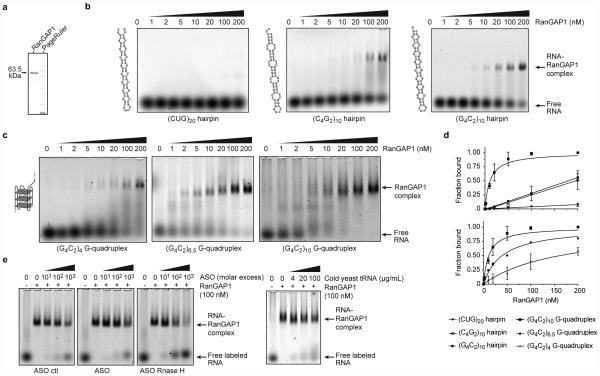
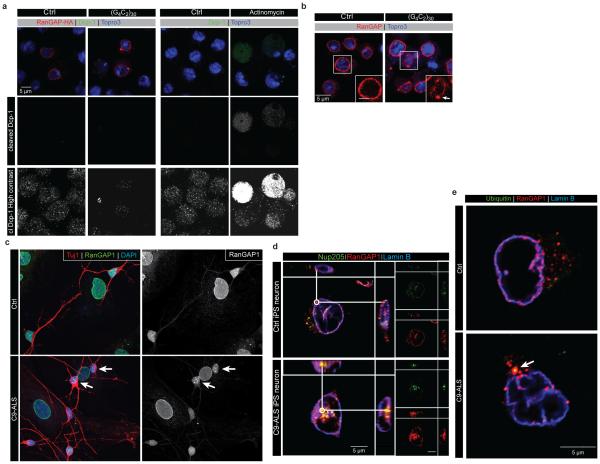
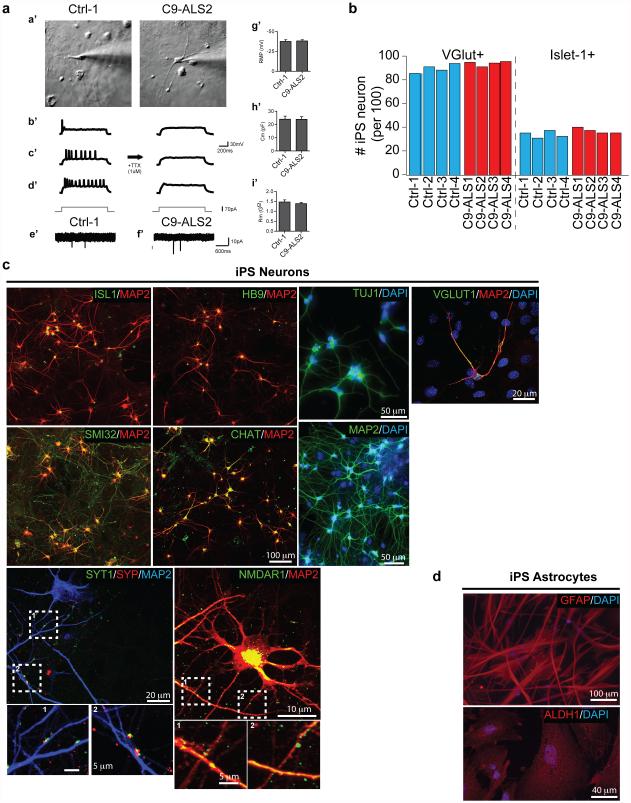
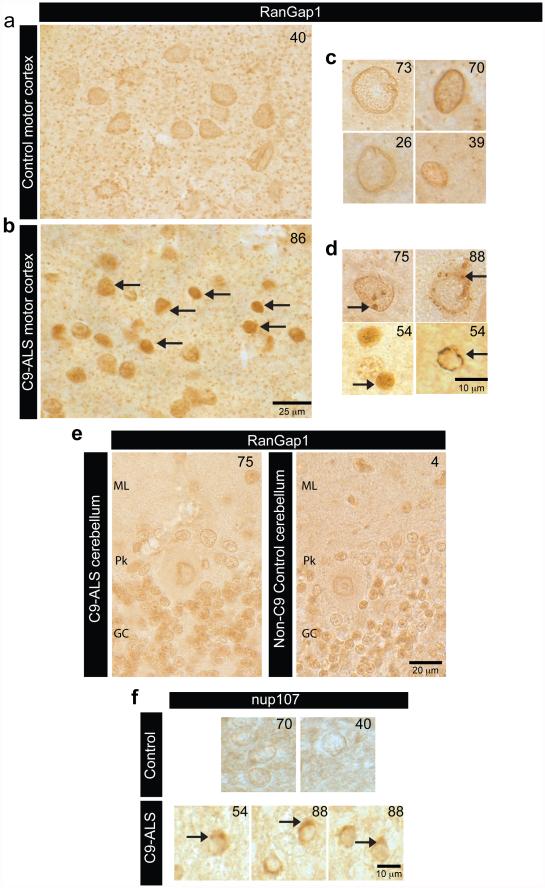
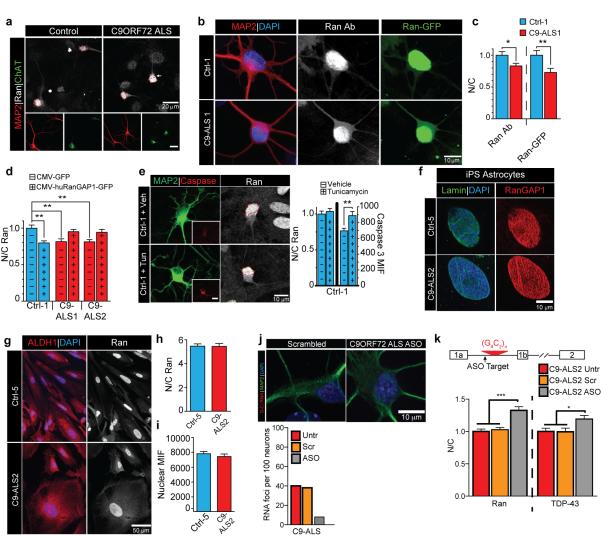
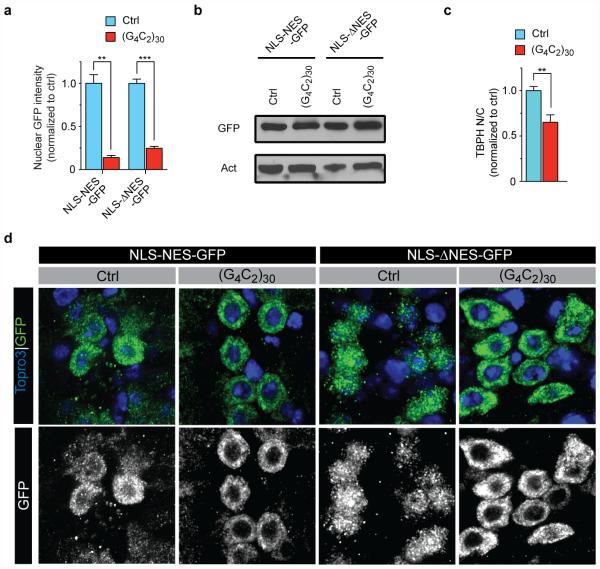
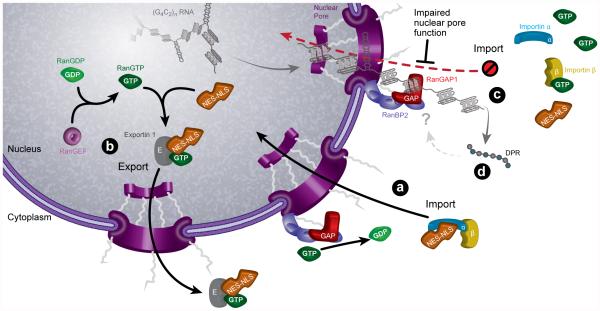
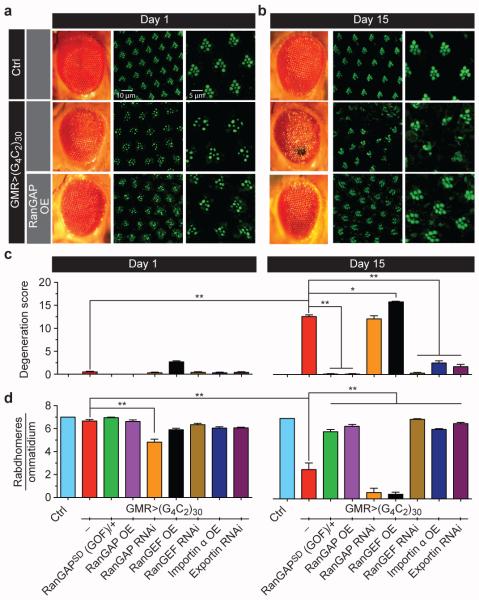
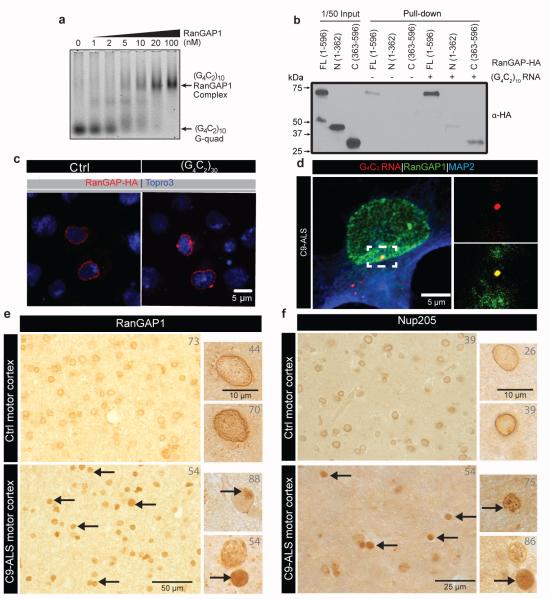
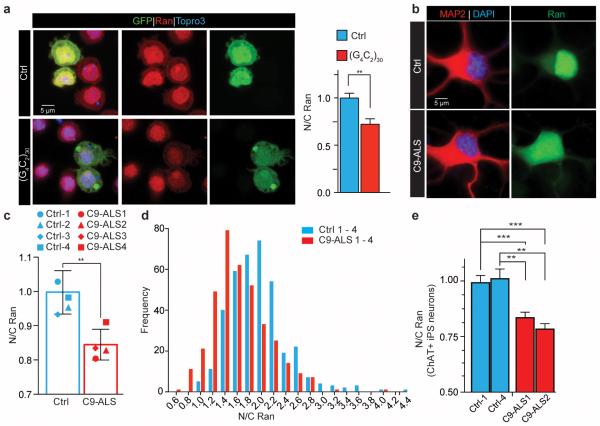
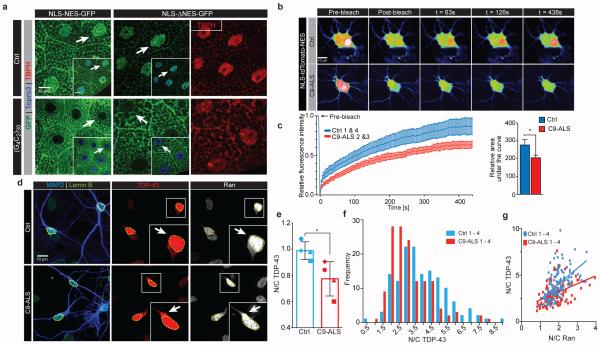
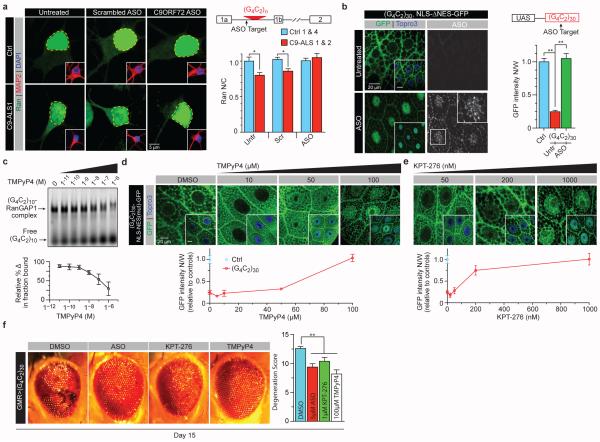
The hexanucleotide repeat expansion (HRE) GGGGCC (G4C2) in C9orf72 is the most common cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Recent studies support an HRE RNA gain-of-function mechanism of neurotoxicity, and we previously identified protein interactors for the G4C2 RNA including RanGAP1. A candidate-based genetic screen in Drosophila expressing 30 G4C2 repeats identified RanGAP (Drosophila orthologue of human RanGAP1), a key regulator of nucleocytoplasmic transport, as a potent suppressor of neurodegeneration. Enhancing nuclear import or suppressing nuclear export of proteins also suppresses neurodegeneration. RanGAP physically interacts with HRE RNA and is mislocalized in HRE-expressing flies, neurons from C9orf72 ALS patient-derived induced pluripotent stem cells (iPSC-derived neurons), and in C9orf72 ALS patient brain tissue. Nuclear import is impaired as a result of HRE expression in the fly model and in C9orf72 iPSC-derived neurons, and these deficits are rescued by small molecules and antisense oligonucleotides targeting the HRE G-quadruplexes. Nucleocytoplasmic transport defects may be a fundamental pathway for ALS and FTD that is amenable to pharmacotherapeutic intervention.
Figures
Comment in
-
Neurodegeneration: Problems at the nuclear pore.Nature. 2015 Sep 3;525(7567):36-7. doi: 10.1038/nature15208. Epub 2015 Aug 26. Nature. 2015. PMID: 26308896 No abstract available.
-
Neurodegenerative disease: C9orf72 repeats compromise nucleocytoplasmic transport.Nat Rev Neurol. 2015 Dec;11(12):670-2. doi: 10.1038/nrneurol.2015.219. Epub 2015 Nov 3. Nat Rev Neurol. 2015. PMID: 26526532 No abstract available.
References
-
- Mori K, et al. The C9orf72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS. Science. 2013;339:1335–1338. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS094239/NS/NINDS NIH HHS/United States
- RC2 NS069395/NS/NINDS NIH HHS/United States
- T32 CA009110/CA/NCI NIH HHS/United States
- R01 NS074324/NS/NINDS NIH HHS/United States
- R01 NS082563/NS/NINDS NIH HHS/United States
- P40OD018537/OD/NIH HHS/United States
- K99 NS091486/NS/NINDS NIH HHS/United States
- U24 NS078736/NS/NINDS NIH HHS/United States
- CA009110/CA/NCI NIH HHS/United States
- R01 NS089616/NS/NINDS NIH HHS/United States
- R01 NS085207/NS/NINDS NIH HHS/United States
- R01 GM060980/GM/NIGMS NIH HHS/United States
- R01NS085207/NS/NINDS NIH HHS/United States
- P40 OD018537/OD/NIH HHS/United States
- R00 NS091486/NS/NINDS NIH HHS/United States
- P01 AG012992/AG/NIA NIH HHS/United States
- NS091046/NS/NINDS NIH HHS/United States
- U54 NS091046/NS/NINDS NIH HHS/United States
- R01 GM084947/GM/NIGMS NIH HHS/United States
- NS089616/NS/NINDS NIH HHS/United States
- R01-GM084947/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
