

Detection of p75NTR Trimers: Implications for Receptor Stoichiometry and Activation
- PMID: 26311773
- PMCID: PMC4549402
- DOI: 10.1523/JNEUROSCI.0591-15.2015
Detection of p75NTR Trimers: Implications for Receptor Stoichiometry and Activation
Abstract
The p75 neurotrophin receptor (p75(NTR)) is a multifunctional receptor that participates in many critical processes in the nervous system, ranging from apoptosis to synaptic plasticity and morphological events. It is a member of the tumor necrosis factor receptor (TNFR) superfamily, whose members undergo trimeric oligomerization. Interestingly, p75(NTR) interacts with dimeric ligands (i.e., proneurotrophins or mature neurotrophins), but several of the intracellular adaptors that mediate p75(NTR) signaling are trimeric (i.e., TNFR-associated factor 6 or TRAF6). Consequently, the active receptor signaling unit remains uncertain. To identify the functional receptor complex, we evaluated its oligomerization in vitro and in mice brain tissues using a combination of biochemical techniques. We found that the most abundant homotypic arrangement for p75(NTR) is a trimer and that monomers and trimers coexist at the cell surface. Interestingly, trimers are not required for ligand-independent or ligand-dependent p75(NTR) activation in a growth cone retraction functional assay. However, monomers are capable of inducing acute morphological effects in neurons. We propose that p75(NTR) activation is regulated by its oligomerization status and its levels of expression. These results indicate that the oligomeric state of p75(NTR) confers differential responses and offers an explanation for the diverse and contradictory actions of this receptor in the nervous system.
Significance statement: The p75 neurotrophin receptor (p75(NTR)) regulates a wide range of cellular functions, including apoptosis, neuronal processes remodeling, and synaptic plasticity. The goal of our work was to inquire whether oligomers of the receptor are required for function. Here we report that p75(NTR) predominantly assembles as a trimer, similar to other tumor necrosis factor receptors. Interestingly, monomers and trimers coexist at the cell surface, but trimers are not required for p75(NTR) activation in a functional assay. However, monomers are capable of inducing acute morphological effects in neurons. Identification of the oligomerization state of p75(NTR) begins to provide insights to the mechanisms of signal initiation of this noncatalytic receptor, as well as to develop therapeutic interventions to diminish its activity.
Keywords: TNF receptor; p75NTR; proNGF; signaling unit; stoichiometry; trimerization.
Copyright © 2015 the authors 0270-6474/15/3511911-10$15.00/0.
Figures
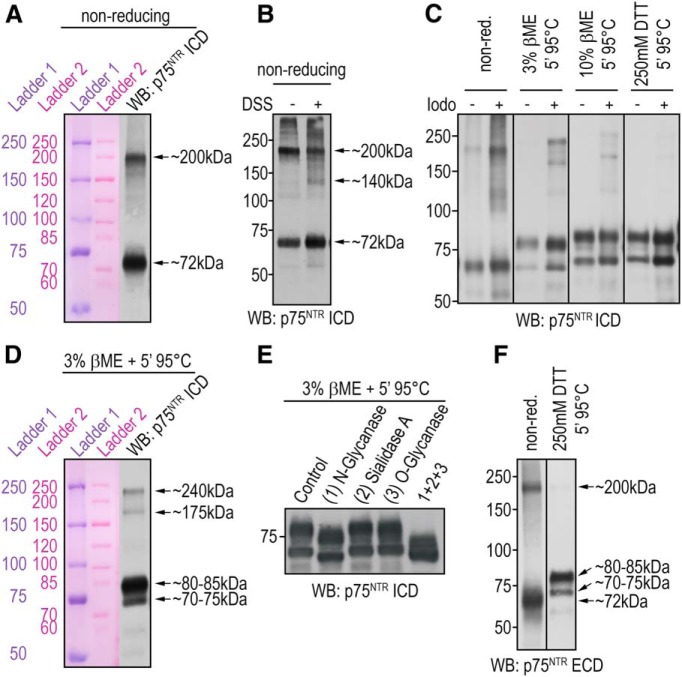
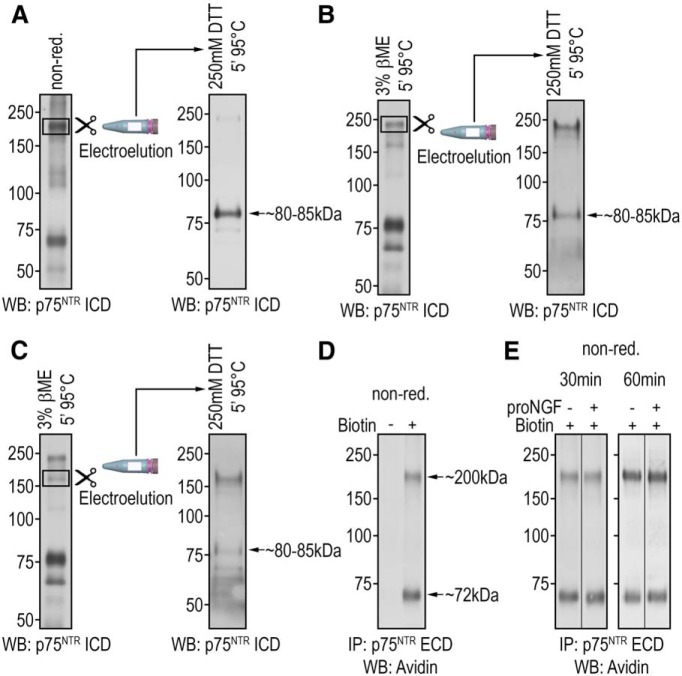
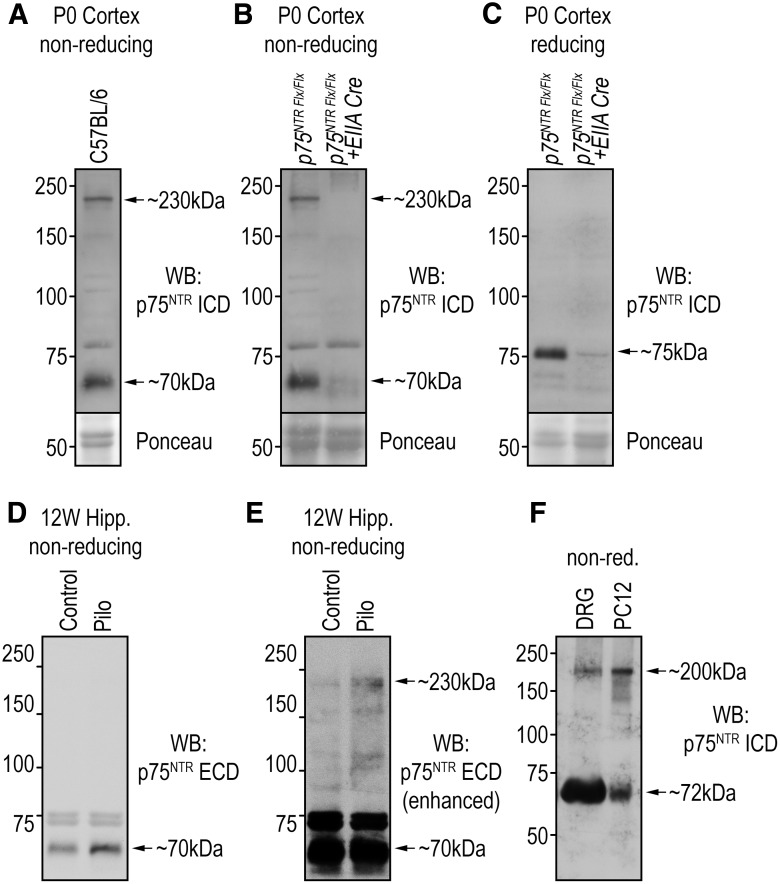
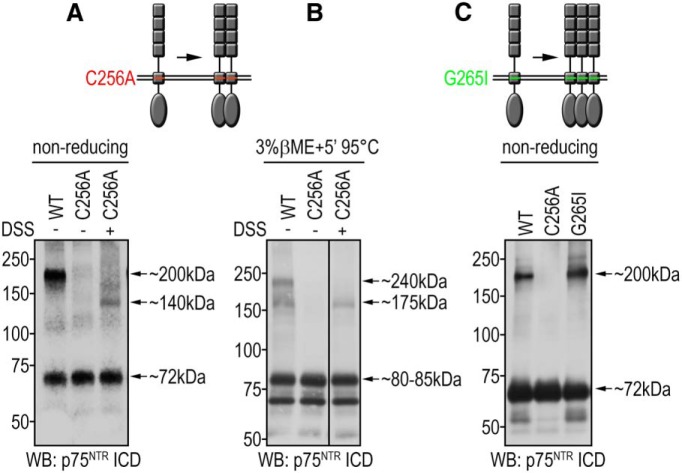
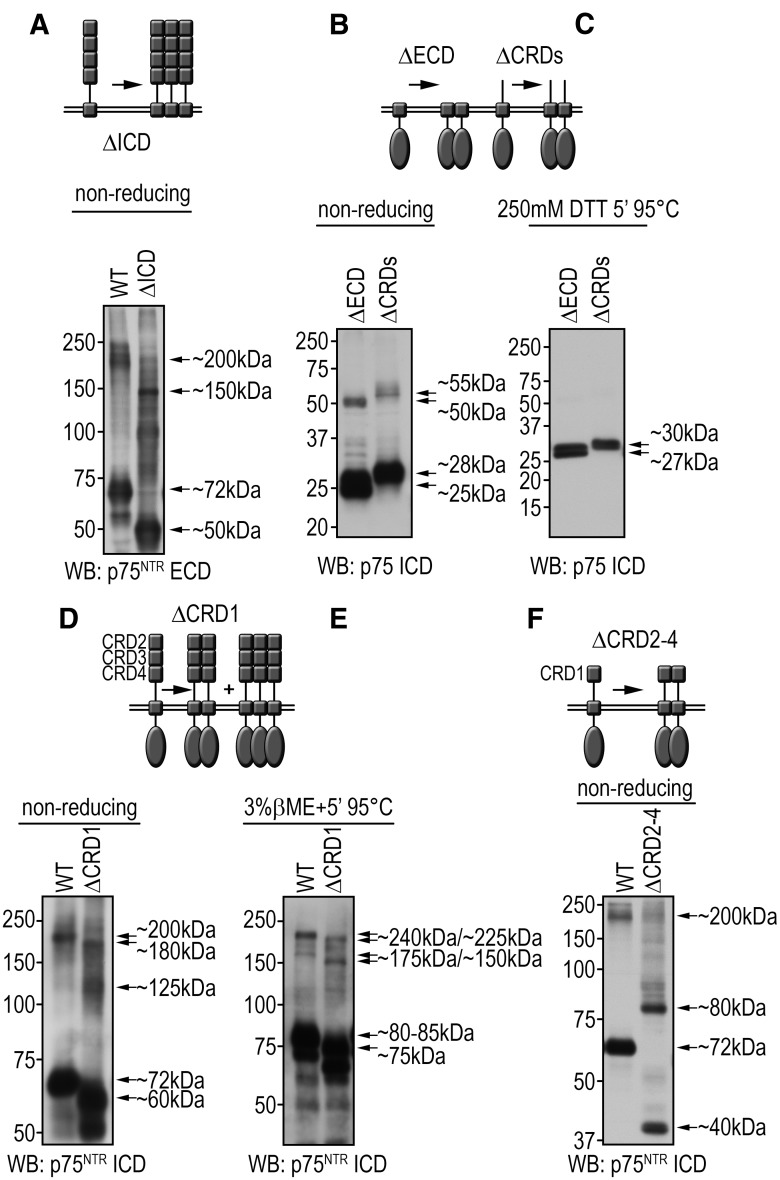
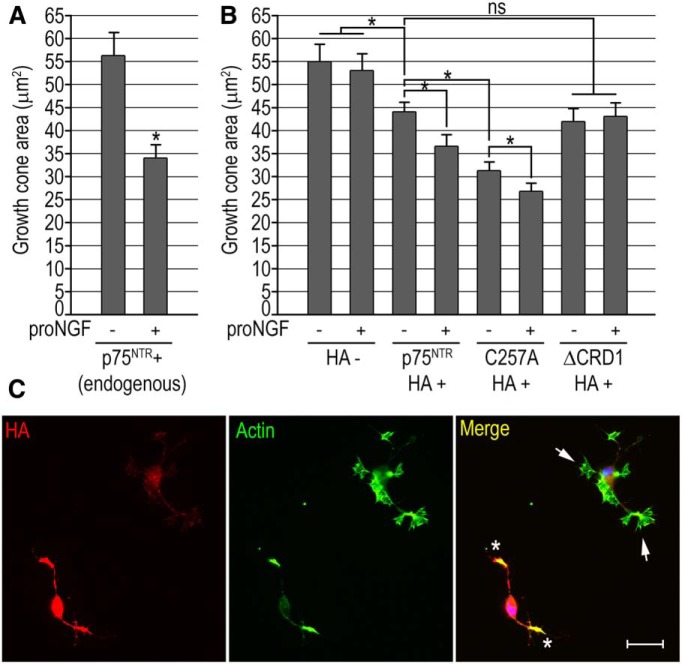
References
Publication types
MeSH terms
Substances
Grants and funding
- NS21072/NS/NINDS NIH HHS/United States
- R01 NS070173/NS/NINDS NIH HHS/United States
- P01 HD023315/HD/NICHD NIH HHS/United States
- R01 NS021072/NS/NINDS NIH HHS/United States
- NS030687/NS/NINDS NIH HHS/United States
- NS070173/NS/NINDS NIH HHS/United States
- 325420/Canadian Institutes of Health Research/Canada
- R01 NS030687/NS/NINDS NIH HHS/United States
- R01 NS064114/NS/NINDS NIH HHS/United States
- AG025970/AG/NIA NIH HHS/United States
- R56 AG025970/AG/NIA NIH HHS/United States
- R56 NS021072/NS/NINDS NIH HHS/United States
- NS064114/NS/NINDS NIH HHS/United States
- HD23315/HD/NICHD NIH HHS/United States
- R01 AG025970/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials