

Pel is a cationic exopolysaccharide that cross-links extracellular DNA in the Pseudomonas aeruginosa biofilm matrix
- PMID: 26311845
- PMCID: PMC4568648
- DOI: 10.1073/pnas.1503058112
Pel is a cationic exopolysaccharide that cross-links extracellular DNA in the Pseudomonas aeruginosa biofilm matrix
Abstract
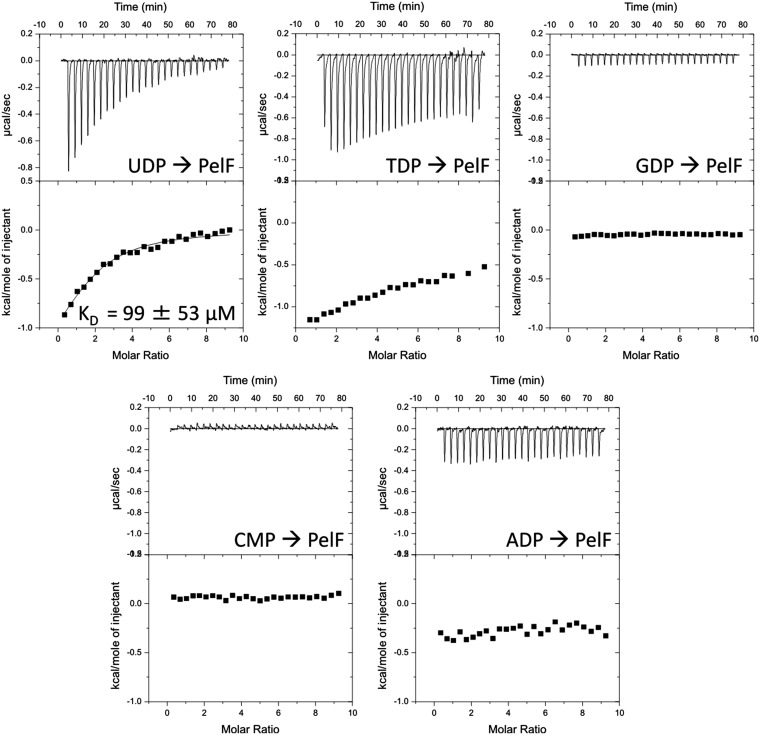
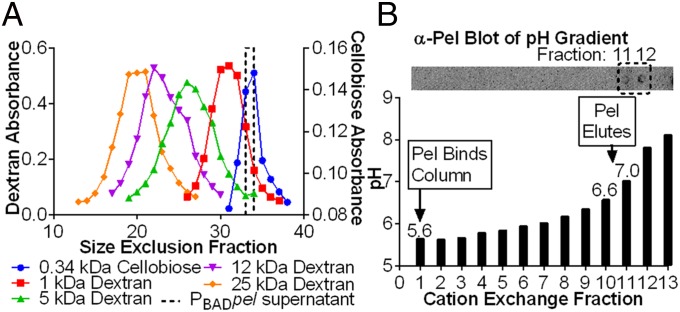
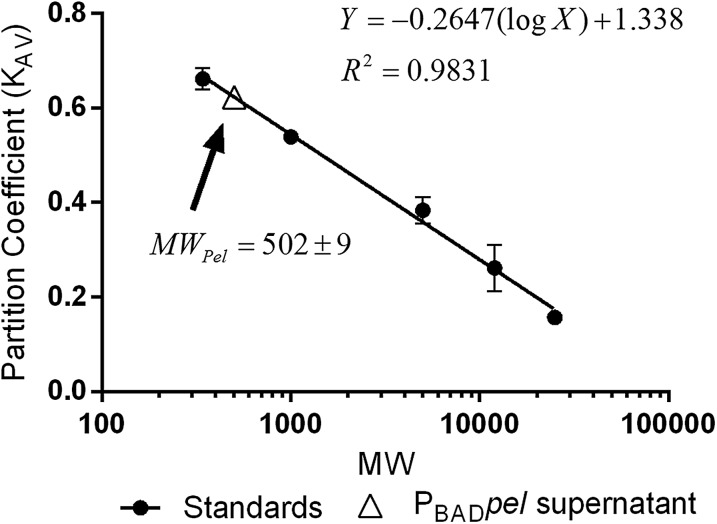
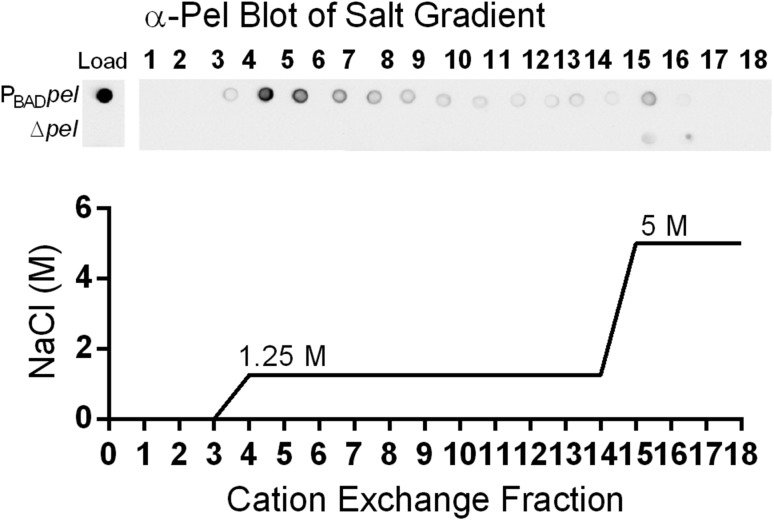
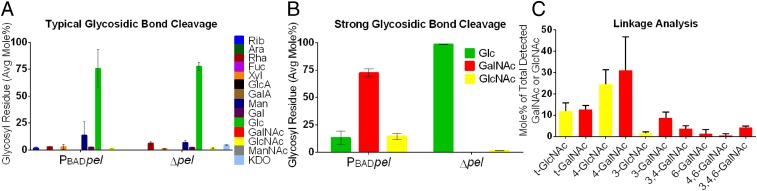
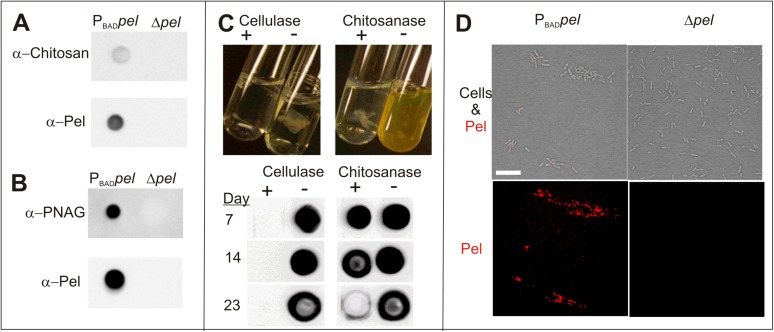
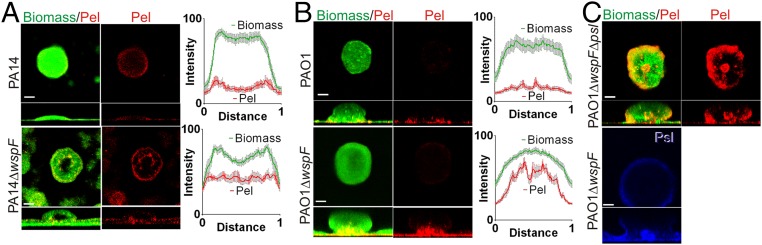
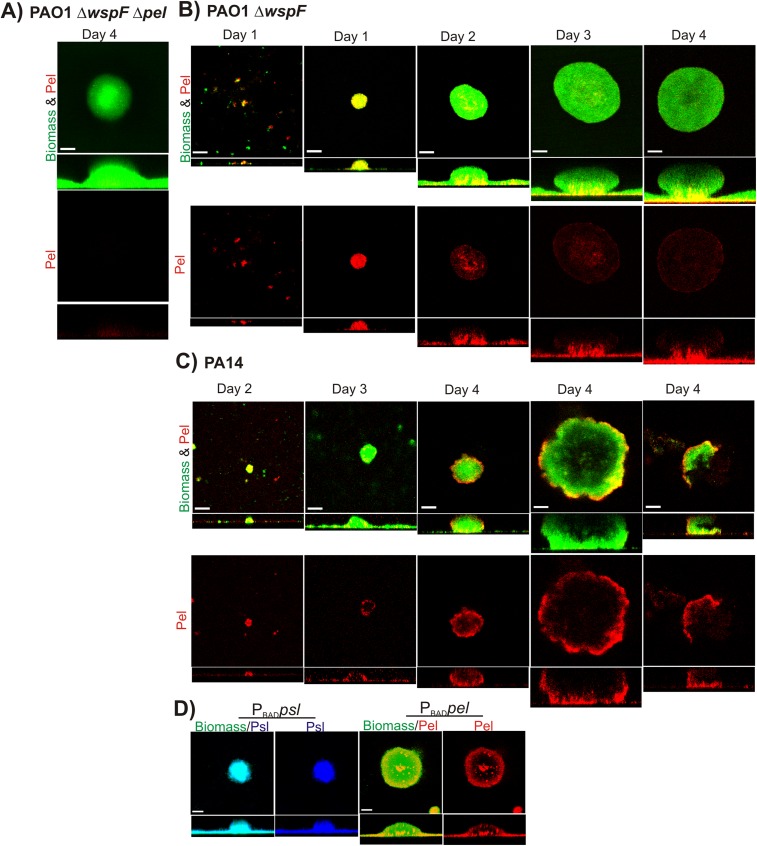
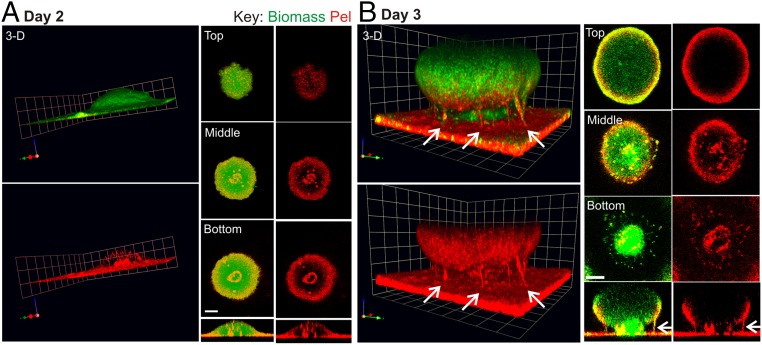
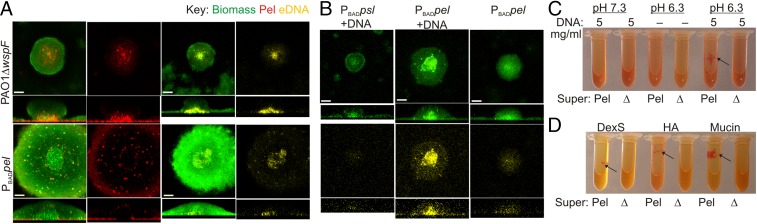
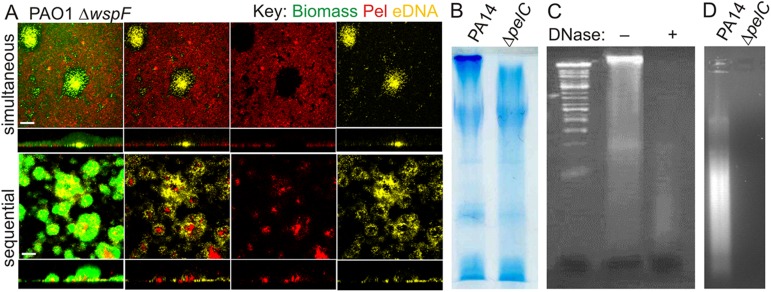
Biofilm formation is a complex, ordered process. In the opportunistic pathogen Pseudomonas aeruginosa, Psl and Pel exopolysaccharides and extracellular DNA (eDNA) serve as structural components of the biofilm matrix. Despite intensive study, Pel's chemical structure and spatial localization within mature biofilms remain unknown. Using specialized carbohydrate chemical analyses, we unexpectedly found that Pel is a positively charged exopolysaccharide composed of partially acetylated 1→4 glycosidic linkages of N-acetylgalactosamine and N-acetylglucosamine. Guided by the knowledge of Pel's sugar composition, we developed a tool for the direct visualization of Pel in biofilms by combining Pel-specific Wisteria floribunda lectin staining with confocal microscopy. The results indicate that Pel cross-links eDNA in the biofilm stalk via ionic interactions. Our data demonstrate that the cationic charge of Pel is distinct from that of other known P. aeruginosa exopolysaccharides and is instrumental in its ability to interact with other key biofilm matrix components.
Keywords: Pel; Psl; biofilms; exopolysaccharide; extracellular DNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: A common cause of persistent infections. Science. 1999;284(5418):1318–1322. - PubMed
-
- Parsek MR, Singh PK. Bacterial biofilms: An emerging link to disease pathogenesis. Annu Rev Microbiol. 2003;57:677–701. - PubMed
-
- Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: From the natural environment to infectious diseases. Nat Rev Microbiol. 2004;2(2):95–108. - PubMed
-
- Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM. Microbial biofilms. Annu Rev Microbiol. 1995;49(1):711–745. - PubMed
-
- Flemming H-C, Wingender J. The biofilm matrix. Nat Rev Microbiol. 2010;8(9):623–633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
