

Mechanism of Binding to Ebola Virus Glycoprotein by the ZMapp, ZMAb, and MB-003 Cocktail Antibodies
- PMID: 26311869
- PMCID: PMC4621129
- DOI: 10.1128/JVI.01490-15
Mechanism of Binding to Ebola Virus Glycoprotein by the ZMapp, ZMAb, and MB-003 Cocktail Antibodies
Abstract
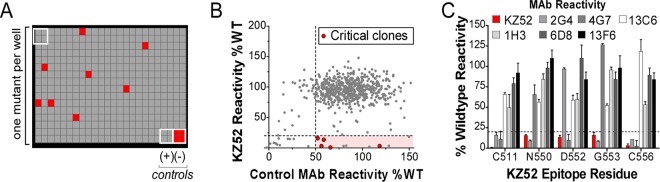
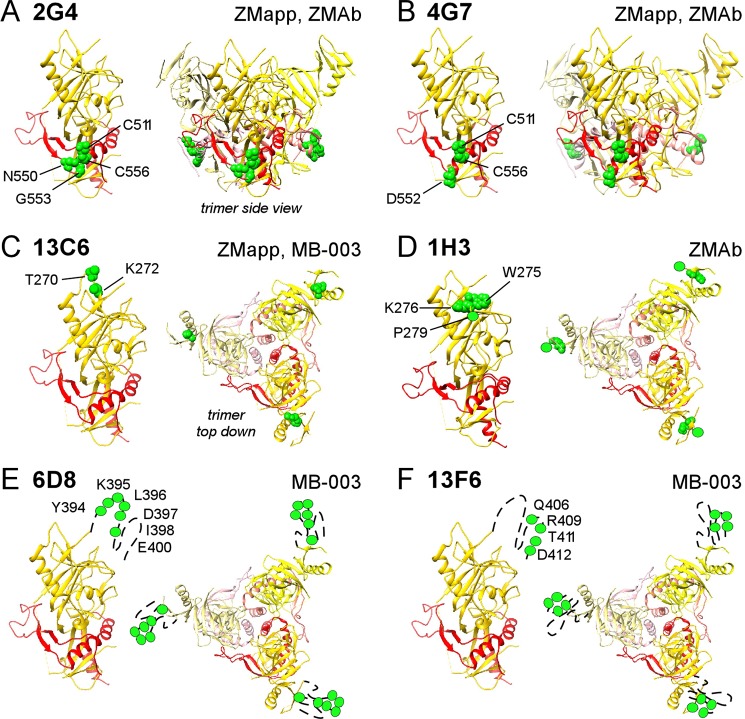
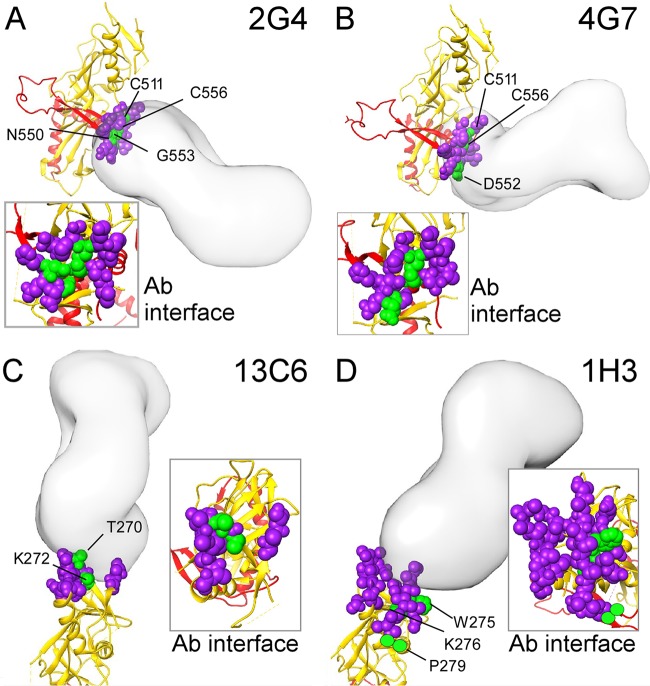
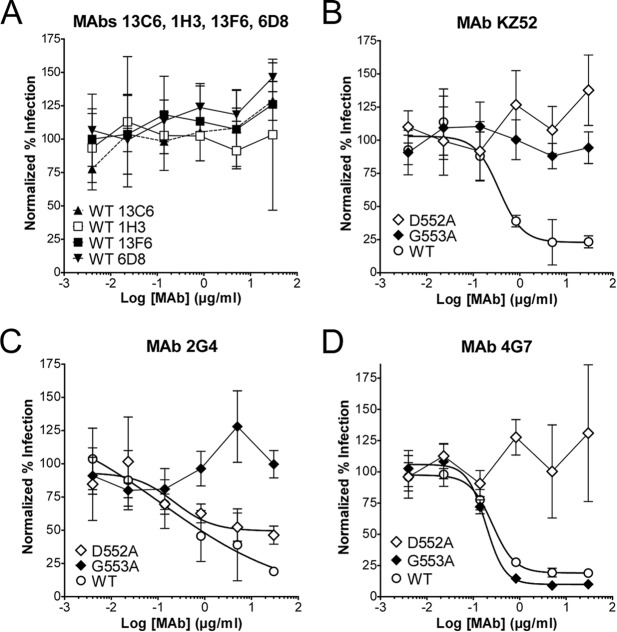
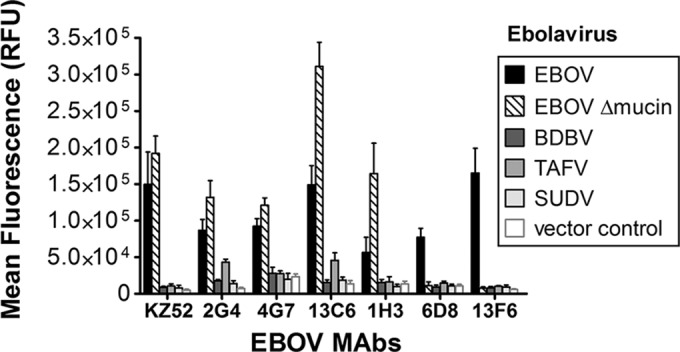
Cocktails of monoclonal antibodies (MAbs) that target the surface glycoprotein (GP) of Ebola virus (EBOV) are effective in nonhuman primate models and have been used under emergency compassionate-treatment protocols in human patients. However, the amino acids that form the detailed binding epitopes for the MAbs in the ZMapp, ZMAb, and the related MB-003 cocktails have yet to be identified. Other binding properties that define how each MAb functionally interacts with GP—such as affinity, epitope conservation, and epitope accessibility—also remain largely unknown. To help define how each MAb interacts with GP, here we used comprehensive alanine-scanning mutagenesis (shotgun mutagenesis), neutralization escape, and whole virion binding to define each MAb's specific epitope, epitope accessibility, epitope conservation, and apparent affinity. Each of the six therapeutic MAbs binds nonidentical epitopes in the GP base, glycan cap, or mucin-like domain. Their apparent affinity, epitope complementarity, and epitope accessibility helps explain why MAbs 4G7 and 13C6 are more protective than 2G4 and 1H3. The mucin-like domain MAbs 6D8 and 13F6 bind with the strongest apparent affinity, helping to explain their effectiveness in vivo despite their inability to neutralize virus.
Importance: Ebola virus disease (EVD) can be caused by four different filovirus family members, including Ebola virus (EBOV), which infected 10 times more people in western Africa over the last year than all previous EVD outbreaks combined, with a number of cases distributed across the globe by travelers. Cocktails of inhibitory monoclonal antibodies (MAbs), such as ZMAb, MB-003, and in particular ZMapp, have demonstrated in animal models some of the most significant therapeutic potential for treating EVD, and in 2014, 15 patients were treated with ZMapp or ZMAb under compassionate-use protocols. Here, we have defined the epitope features for the most important therapeutic MAbs against EBOV developed to date. Defining the epitopes and binding characteristics for these MAbs, as well as the commonly used reference MAb KZ52, helps explain their breadth of reactivity against different ebolavirus species, predict viral evasion against these MAbs, and design new cocktails of MAbs with improved complementarity.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- World Health Organization. 2015. Ebola situation report—8 July 2015. World Health Organization, Geneva, Switzerland: http://apps.who.int/ebola/current-situation/ebola-situation-report-8-jul....
-
- Kuhn JH, Andersen KG, Bao Y, Bavari S, Becker S, Bennett RS, Bergman NH, Blinkova O, Bradfute S, Brister JR, Bukreyev A, Chandran K, Chepurnov AA, Davey RA, Dietzgen RG, Doggett NA, Dolnik O, Dye JM, Enterlein S, Fenimore PW, Formenty P, Freiberg AN, Garry RF, Garza NL, Gire SK, Gonzalez JP, Griffiths A, Happi CT, Hensley LE, Herbert AS, Hevey MC, Hoenen T, Honko AN, Ignatyev GM, Jahrling PB, Johnson JC, Johnson KM, Kindrachuk J, Klenk HD, Kobinger G, Kochel TJ, Lackemeyer MG, Lackner DF, Leroy EM, Lever MS, Muhlberger E, Netesov SV, Olinger GG, Omilabu SA, Palacios G, et al. 2014. Filovirus RefSeq entries: evaluation and selection of filovirus type variants, type sequences, and names. Viruses 6:3663–3682. doi: 10.3390/v6093663. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
