

Differences in Processing Determinants of Nonstructural Polyprotein and in the Sequence of Nonstructural Protein 3 Affect Neurovirulence of Semliki Forest Virus
- PMID: 26311875
- PMCID: PMC4621116
- DOI: 10.1128/JVI.01186-15
Differences in Processing Determinants of Nonstructural Polyprotein and in the Sequence of Nonstructural Protein 3 Affect Neurovirulence of Semliki Forest Virus
Abstract
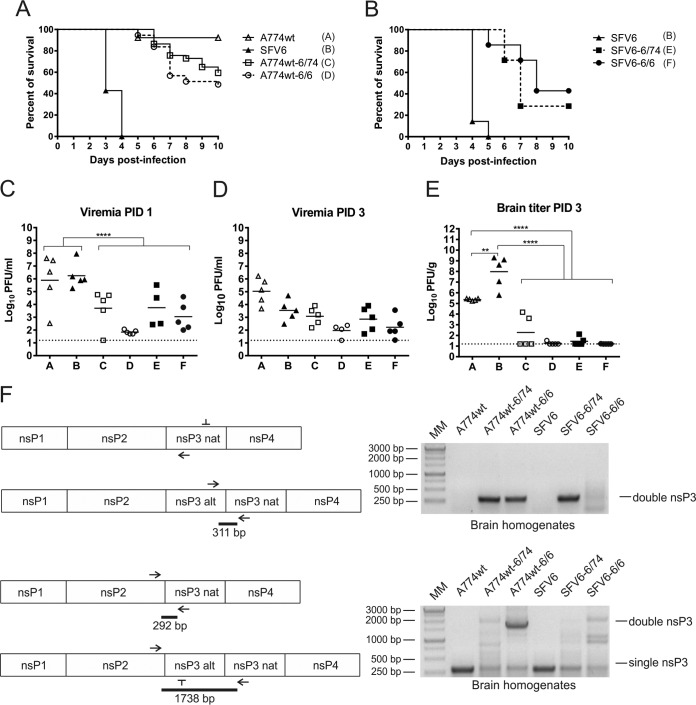
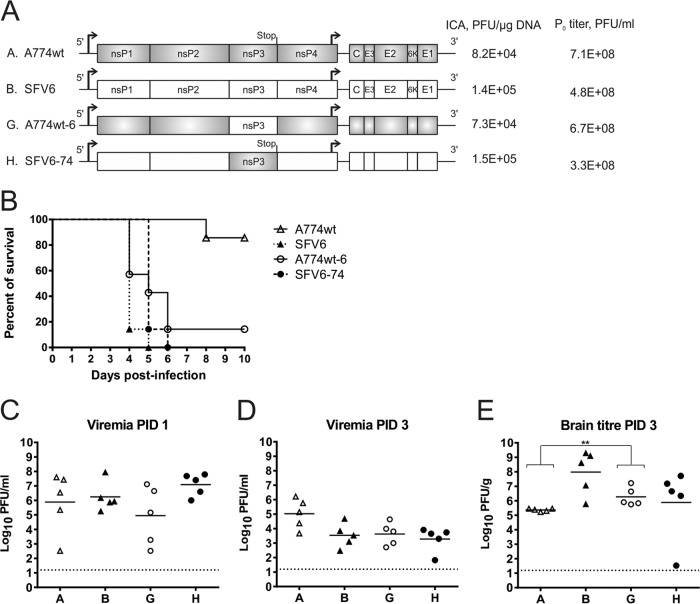
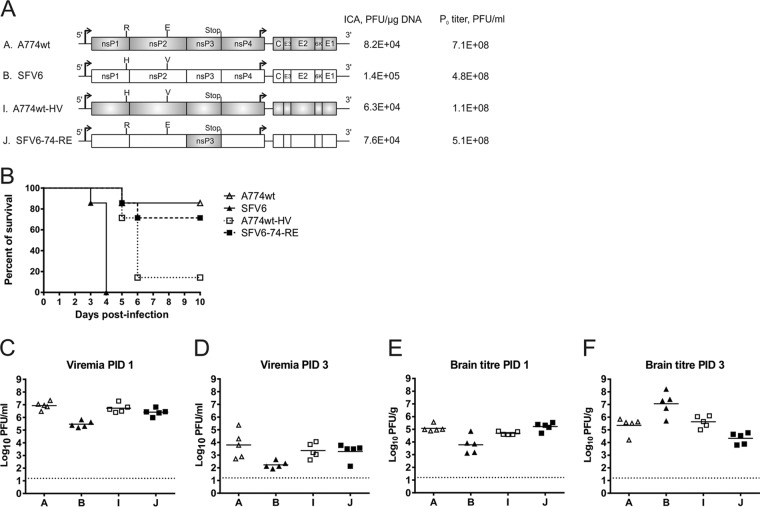
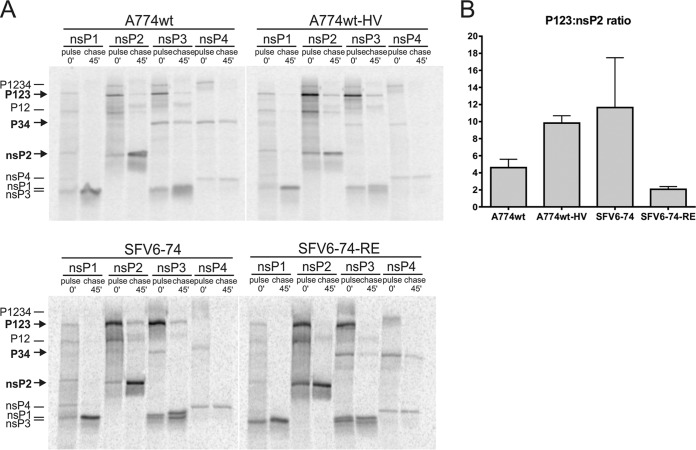
The A7(74) strain of Semliki Forest virus (SFV; genus Alphavirus) is avirulent in adult mice, while the L10 strain is virulent in mice of all ages. It has been previously demonstrated that this phenotypic difference is associated with nonstructural protein 3 (nsP3). Consensus clones of L10 (designated SFV6) and A7(74) (designated A774wt) were used to construct a panel of recombinant viruses. The insertion of nsP3 from A774wt into the SFV6 backbone had a minor effect on the virulence of the resulting recombinant virus. Conversely, insertion of nsP3 from SFV6 into the A774wt backbone or replacement of A774wt nsP3 with two copies of nsP3 from SFV6 resulted in virulent viruses. Unexpectedly, duplication of nsP3-encoding sequences also resulted in elevated levels of nsP4, revealing that nsP3 is involved in the stabilization of nsP4. Interestingly, replacement of nsP3 of SFV6 with that of A774wt resulted in a virulent virus; the virulence of this recombinant was strongly reduced by functionally coupled substitutions for amino acid residues 534 (P4 position of the cleavage site between nsP1 and nsP2) and 1052 (S4 subsite residue of nsP2 protease) in the nonstructural polyprotein. Pulse-chase experiments revealed that A774wt and avirulent recombinant virus were characterized by increased processing speed of the cleavage site between nsP1 and nsP2. A His534-to-Arg substitution specifically activated this cleavage, while a Val1052-to-Glu substitution compensated for this effect by reducing the basal protease activity of nsP2. These findings provide a link between nonstructural polyprotein processing and the virulence of SFV.
Importance: SFV infection of mice provides a well-characterized model to study viral encephalitis. SFV also serves as a model for studies of alphavirus molecular biology and host-pathogen interactions. Thus far, the genetic basis of different properties of SFV strains has been studied using molecular clones, which often contain mistakes originating from standard cDNA synthesis and cloning procedures. Here, for the first time, consensus clones of SFV strains were used to map virulence determinants. Existing data on the importance of nsP3 for virulent phenotypes were confirmed, another determinant of neurovirulence and its molecular basis was characterized, and a novel function of nsP3 was identified. These findings provide links between the molecular biology of SFV and its biological properties and significantly increase our understanding of the basis of alphavirus-induced pathology. In addition, the usefulness of consensus clones as tools for studies of alphaviruses was demonstrated.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures

Similar articles
-
Regulation of Semliki Forest virus RNA replication: a model for the control of alphavirus pathogenesis in invertebrate hosts.Virology. 2004 May 20;323(1):153-63. doi: 10.1016/j.virol.2004.03.009. Virology. 2004. PMID: 15165827
-
Amino acid mutations in the replicase protein nsP3 of Semliki Forest virus cumulatively affect neurovirulence.J Gen Virol. 2003 Jun;84(Pt 6):1525-1533. doi: 10.1099/vir.0.18936-0. J Gen Virol. 2003. PMID: 12771422
-
Semliki Forest Virus Chimeras with Functional Replicase Modules from Related Alphaviruses Survive by Adaptive Mutations in Functionally Important Hot Spots.J Virol. 2021 Sep 27;95(20):e0097321. doi: 10.1128/JVI.00973-21. Epub 2021 Jul 28. J Virol. 2021. PMID: 34319778 Free PMC article.
-
Molecular determinants of alphavirus neuropathogenesis in mice.J Gen Virol. 2016 Jun;97(6):1283-1296. doi: 10.1099/jgv.0.000467. Epub 2016 Mar 30. J Gen Virol. 2016. PMID: 27028153 Review.
-
The Putative Roles and Functions of Indel, Repetition and Duplication Events in Alphavirus Non-Structural Protein 3 Hypervariable Domain (nsP3 HVD) in Evolution, Viability and Re-Emergence.Viruses. 2021 May 28;13(6):1021. doi: 10.3390/v13061021. Viruses. 2021. PMID: 34071712 Free PMC article. Review.
Cited by
-
ADP-ribosyl-binding and hydrolase activities of the alphavirus nsP3 macrodomain are critical for initiation of virus replication.Proc Natl Acad Sci U S A. 2018 Oct 30;115(44):E10457-E10466. doi: 10.1073/pnas.1812130115. Epub 2018 Oct 15. Proc Natl Acad Sci U S A. 2018. PMID: 30322911 Free PMC article.
-
Attenuated Getah virus confers protection against multiple arthritogenic alphaviruses.PLoS Pathog. 2024 Nov 18;20(11):e1012700. doi: 10.1371/journal.ppat.1012700. eCollection 2024 Nov. PLoS Pathog. 2024. PMID: 39556619 Free PMC article.
-
ADP-ribosylhydrolase activity of Chikungunya virus macrodomain is critical for virus replication and virulence.Proc Natl Acad Sci U S A. 2017 Feb 14;114(7):1666-1671. doi: 10.1073/pnas.1621485114. Epub 2017 Jan 31. Proc Natl Acad Sci U S A. 2017. PMID: 28143925 Free PMC article.
-
Decreased Virulence of Ross River Virus Harboring a Mutation in the First Cleavage Site of Nonstructural Polyprotein Is Caused by a Novel Mechanism Leading to Increased Production of Interferon-Inducing RNAs.mBio. 2018 Aug 21;9(4):e00044-18. doi: 10.1128/mBio.00044-18. mBio. 2018. PMID: 30131356 Free PMC article.
-
Characterization of the Zika virus induced small RNA response in Aedes aegypti cells.PLoS Negl Trop Dis. 2017 Oct 17;11(10):e0006010. doi: 10.1371/journal.pntd.0006010. eCollection 2017 Oct. PLoS Negl Trop Dis. 2017. PMID: 29040304 Free PMC article.
References
-
- Fazakerley JK, Cotterill CL, Lee G, Graham A. 2006. Virus tropism, distribution, persistence and pathology in the corpus callosum of the Semliki Forest virus-infected mouse brain: a novel system to study virus-oligodendrocyte interactions. Neuropathol Appl Neurobiol 32:397–409. doi:10.1111/j.1365-2990.2006.00739.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
