

Differential Requirements for L-Citrulline and L-Arginine during Antimycobacterial Macrophage Activity
- PMID: 26311904
- PMCID: PMC6432794
- DOI: 10.4049/jimmunol.1500800
Differential Requirements for L-Citrulline and L-Arginine during Antimycobacterial Macrophage Activity
Abstract
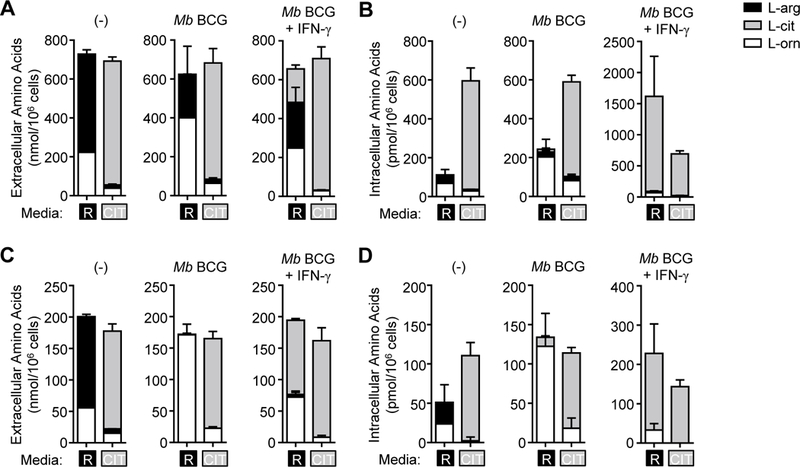
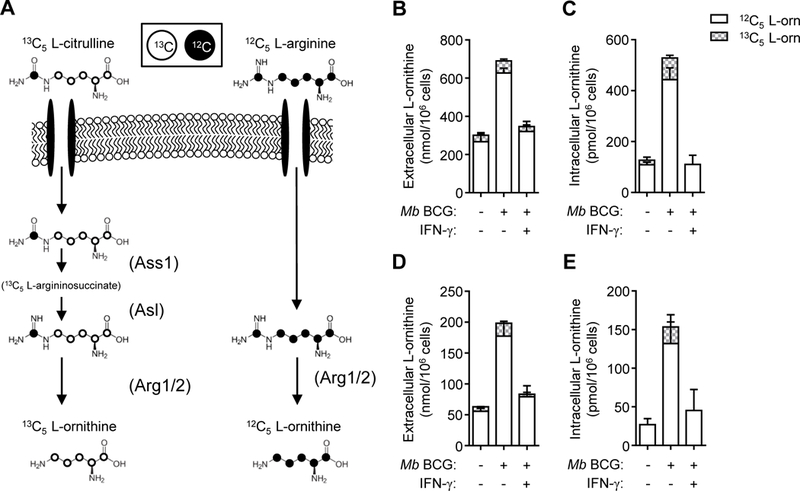
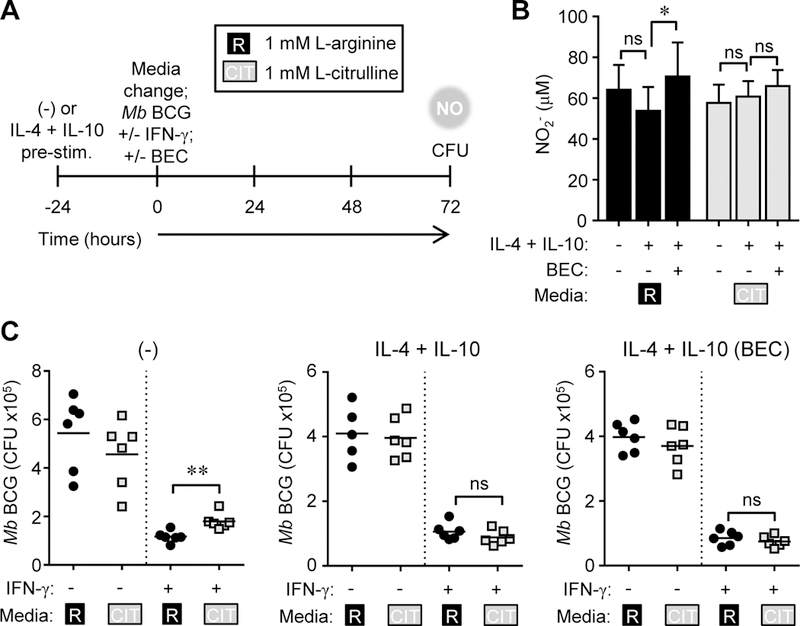
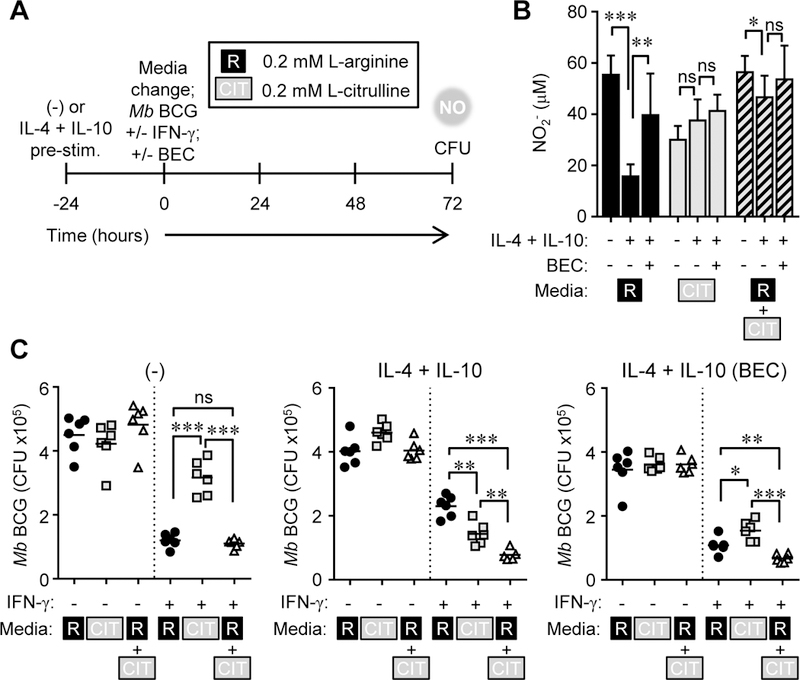
Microbicidal NO production is reliant on inducible NO synthase-mediated L-arginine metabolism in macrophages (MΦs). However, L-arginine supply can be restricted by arginase activity, resulting in inefficient NO output and inhibition of antimicrobial MΦ function. MΦs circumvent this by converting L-citrulline to L-arginine, thereby resupplying substrate for NO production. In this article, we define the metabolic signature of mycobacteria-infected murine MΦs supplied L-arginine, L-citrulline, or both amino acids. Using liquid chromatography-tandem mass spectrometry, we determined that L-arginine synthesized from L-citrulline was less effective as a substrate for arginase-mediated L-ornithine production compared with L-arginine directly imported from the extracellular milieu. Following Mycobacterium bovis bacillus Calmette-Guérin infection and costimulation with IFN-γ, we observed that MΦ arginase activity did not inhibit production of NO derived from L-citrulline, contrary to NO inhibition witnessed when MΦs were cultured in L-arginine. Furthermore, we found that arginase-expressing MΦs preferred L-citrulline over L-arginine for the promotion of antimycobacterial activity. We expect that defining the consequences of L-citrulline metabolism in MΦs will provide novel approaches for enhancing immunity, especially in the context of mycobacterial disease.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures




References
-
- Garcia I, Guler R, Vesin D, Olleros ML, Vassalli P, Chvatchko Y, Jacobs M, and Ryffel B. 2000. Lethal Mycobacterium Bovis Bacillus Calmette Guerin Infection in Nitric Oxide Synthase 2-Deficient Mice: Cell-Mediated Immunity Requires Nitric Oxide Synthase 2. Lab Invest 80: 1385–1397. - PubMed
-
- Gomez LM, Anaya JM, Vilchez JR, Cadena J, Hinojosa R, Velez L, Lopez- Nevot MA, and Martin J. 2007. A Polymorphism in the Inducible Nitric Oxide Synthase Gene Is Associated with Tuberculosis. Tuberculosis (Edinb) 87: 288–294. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
