

Proteomic analysis of pRb loss highlights a signature of decreased mitochondrial oxidative phosphorylation
- PMID: 26314710
- PMCID: PMC4573859
- DOI: 10.1101/gad.264127.115
Proteomic analysis of pRb loss highlights a signature of decreased mitochondrial oxidative phosphorylation
Abstract
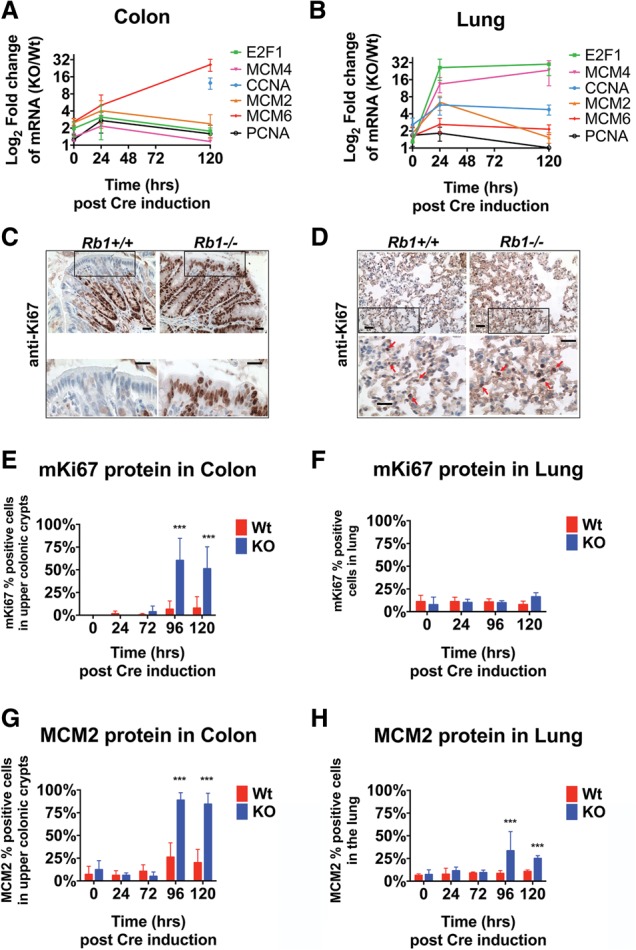
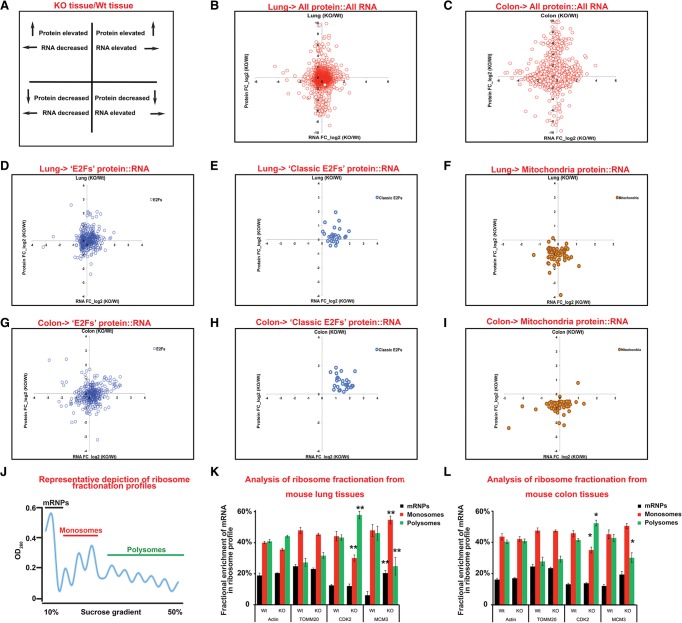
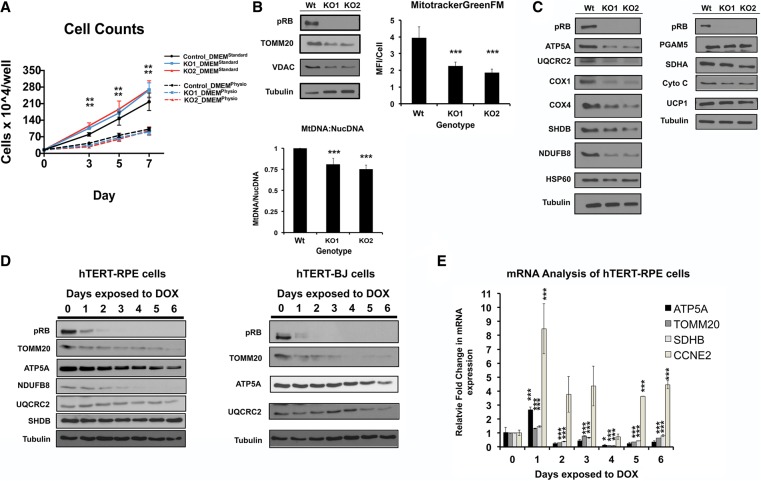
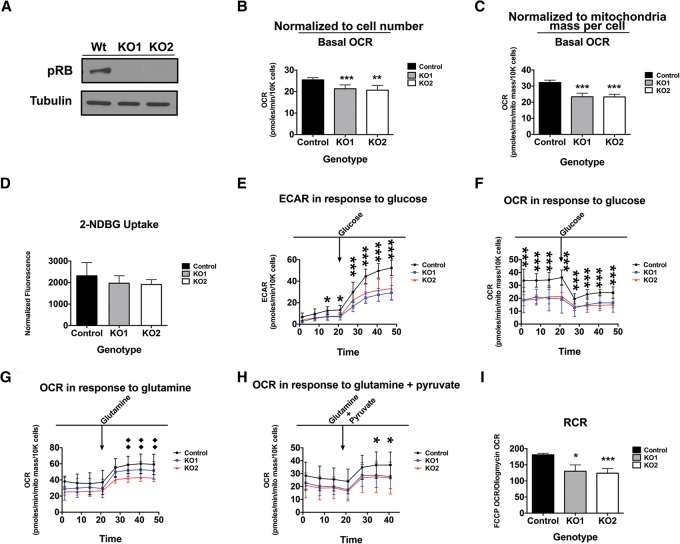
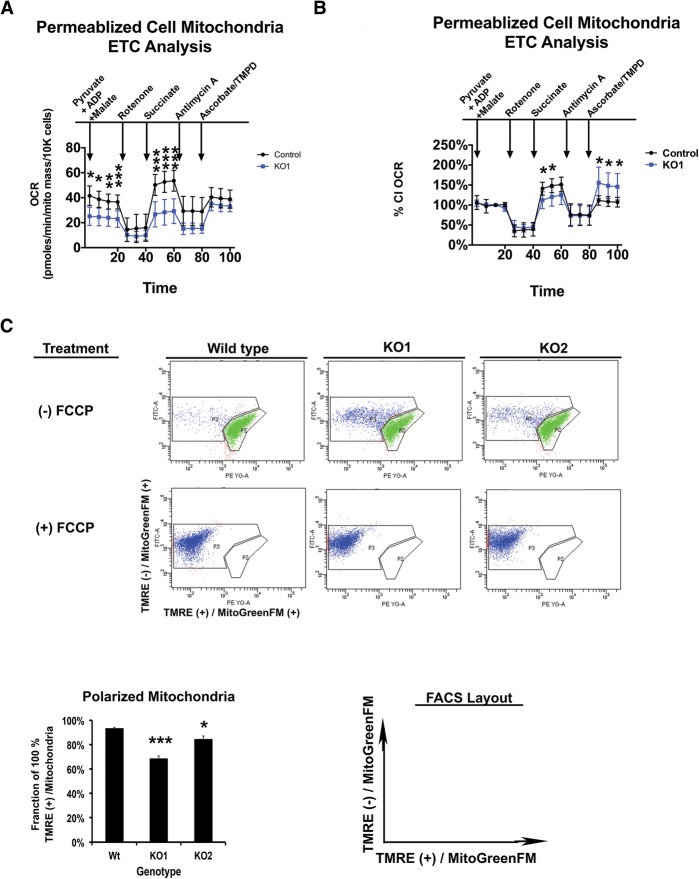
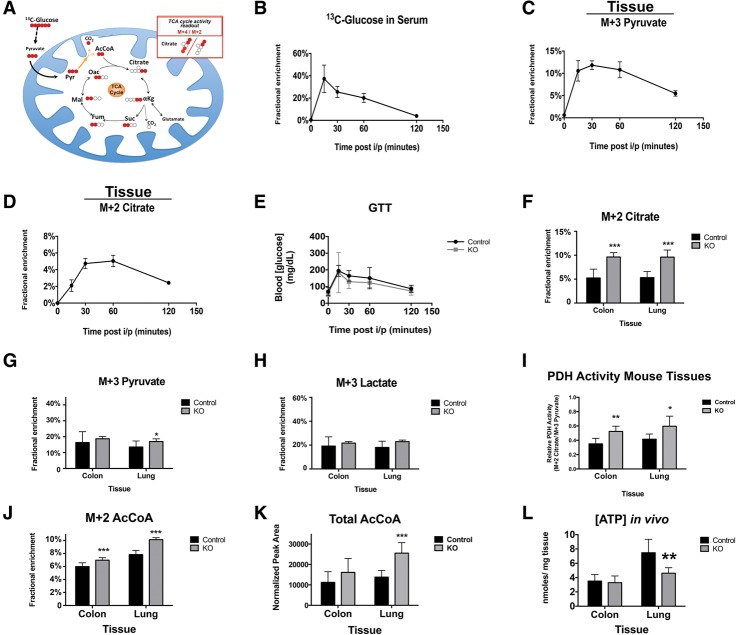
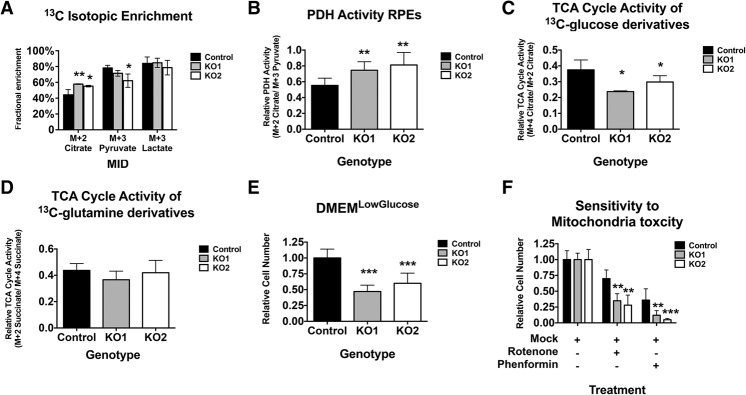
The retinoblastoma tumor suppressor (pRb) protein associates with chromatin and regulates gene expression. Numerous studies have identified Rb-dependent RNA signatures, but the proteomic effects of Rb loss are largely unexplored. We acutely ablated Rb in adult mice and conducted a quantitative analysis of RNA and proteomic changes in the colon and lungs, where Rb(KO) was sufficient or insufficient to induce ectopic proliferation, respectively. As expected, Rb(KO) caused similar increases in classic pRb/E2F-regulated transcripts in both tissues, but, unexpectedly, their protein products increased only in the colon, consistent with its increased proliferative index. Thus, these protein changes induced by Rb loss are coupled with proliferation but uncoupled from transcription. The proteomic changes in common between Rb(KO) tissues showed a striking decrease in proteins with mitochondrial functions. Accordingly, RB1 inactivation in human cells decreased both mitochondrial mass and oxidative phosphorylation (OXPHOS) function. RB(KO) cells showed decreased mitochondrial respiratory capacity and the accumulation of hypopolarized mitochondria. Additionally, RB/Rb loss altered mitochondrial pyruvate oxidation from (13)C-glucose through the TCA cycle in mouse tissues and cultured cells. Consequently, RB(KO) cells have an enhanced sensitivity to mitochondrial stress conditions. In summary, proteomic analyses provide a new perspective on Rb/RB1 mutation, highlighting the importance of pRb for mitochondrial function and suggesting vulnerabilities for treatment.
Keywords: 13C-glucose; OXPHOS; metabolism; mitochondria; pRB; proteomics.
© 2015 Nicolay et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous