

Roles of Integrins and Intracellular Molecules in the Migration and Neuritogenesis of Fetal Cortical Neurons: MEK Regulates Only the Neuritogenesis
- PMID: 26317102
- PMCID: PMC4437273
- DOI: 10.1155/2013/859257
Roles of Integrins and Intracellular Molecules in the Migration and Neuritogenesis of Fetal Cortical Neurons: MEK Regulates Only the Neuritogenesis
Abstract
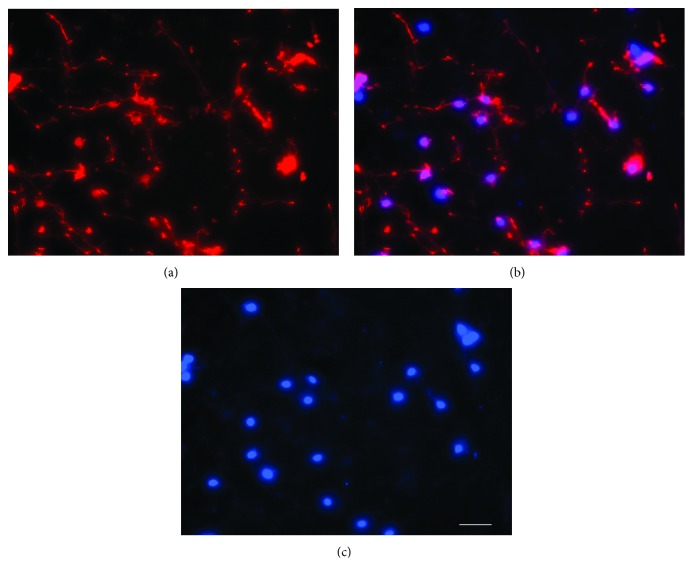
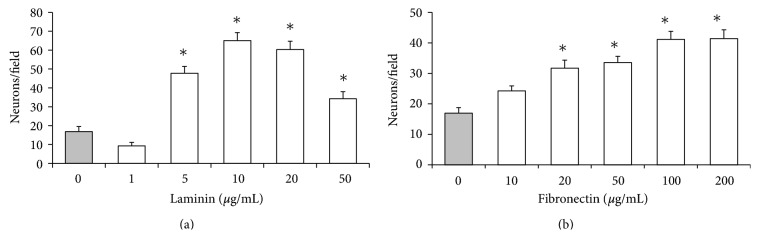
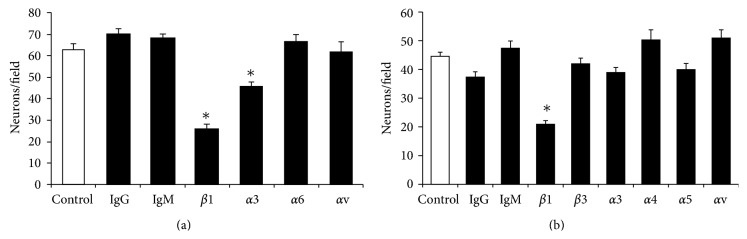
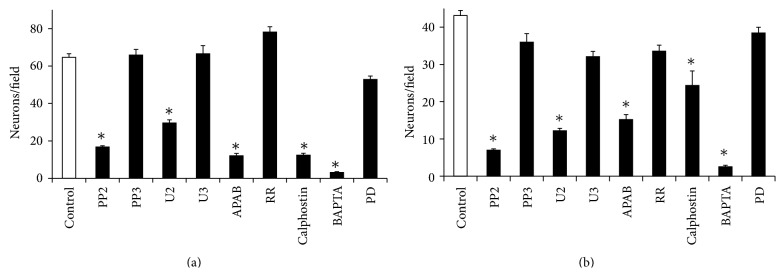
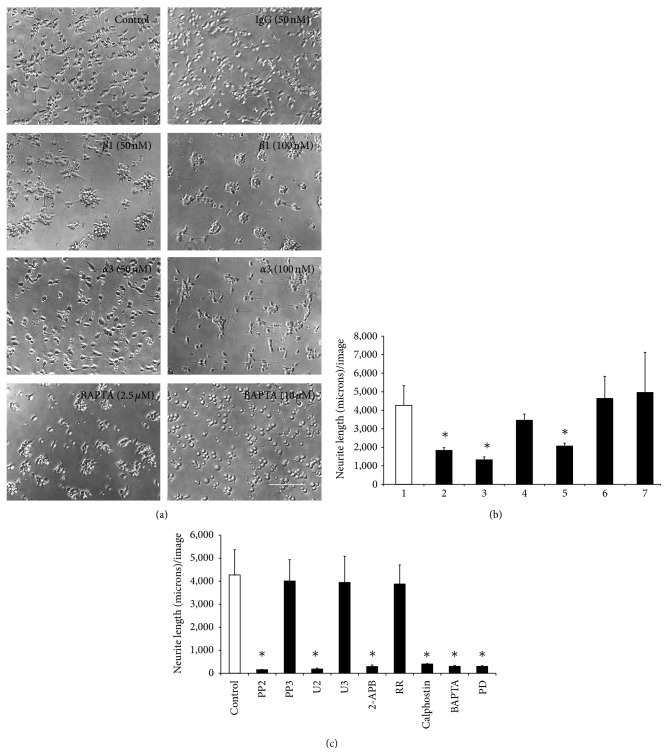
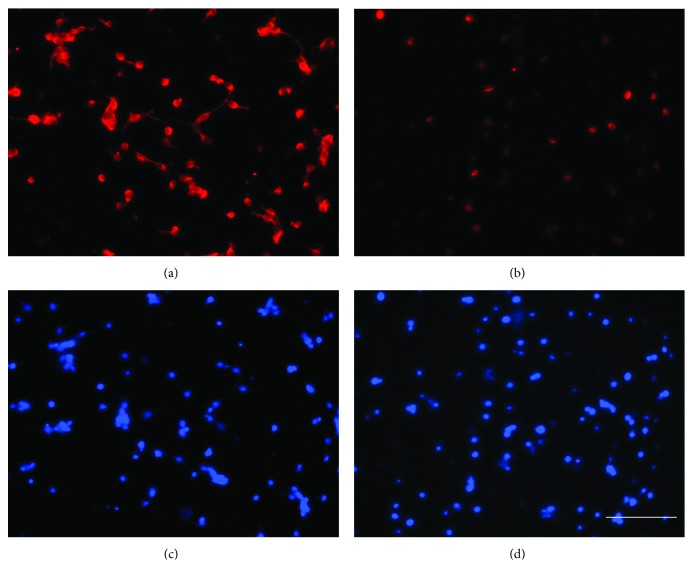
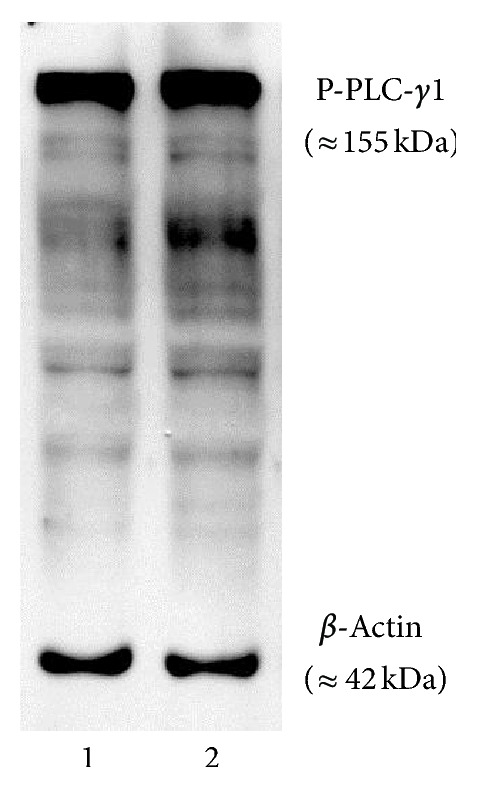
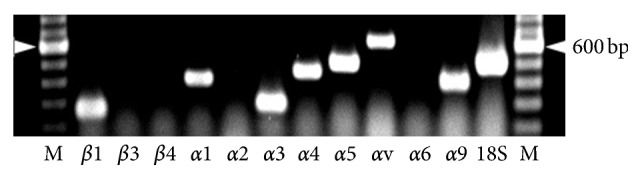
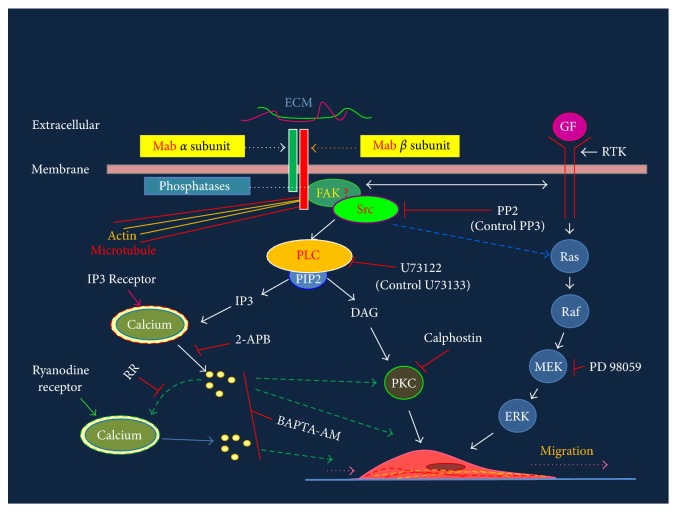
The roles of integrin subunits and intracellular molecules in regulating the migration and neuritogenesis of neurons isolated from 16.5 gestation days rat fetal cortices were examined using in vitro assays. Results showed that laminin supported the migration of fetal cortical neurons better than fibronectin and that the fetal cortical neurons migrated on laminin using β1 and α3 integrin subunits which make up the α3β1 integrin receptor. On fibronectin, the migration was mediated by β1 integrin subunit. Perturbation of src kinase, phospholipase C, or protein kinase C activity, inhibition of IP3 receptor mediated calcium release, or chelation of intracellular calcium inhibited both migration and neuritogenesis, whereas inhibition of growth factor signaling via MEK inhibited only the neuritogenesis. The detection of α1 and α9 transcripts suggested that the migration of fetal cortical neurons may also be mediated by α1β1 and α9β1 integrin receptors. Results showed that calcium may regulate migration and neuritogenesis by maintaining optimum levels of microtubules in the fetal cortical neurons. It is concluded that the fetal cortical neurons are fully equipped with the integrin signaling cascade required for their migration and neuritogenesis, whereas crosstalk between the integrin and growth-factor signaling regulate only the neuritogenesis.
Figures

References
-
- Anton E. S., Kreidberg J. A., Rakic P. Distinct functions of α3 and α(v) integrin receptors in neuronal migration and laminar organization of the cerebral cortex. Neuron. 1999;22(2):277–289. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
