

Bile Salts Modulate the Mucin-Activated Type VI Secretion System of Pandemic Vibrio cholerae
- PMID: 26317760
- PMCID: PMC4552747
- DOI: 10.1371/journal.pntd.0004031
Bile Salts Modulate the Mucin-Activated Type VI Secretion System of Pandemic Vibrio cholerae
Abstract
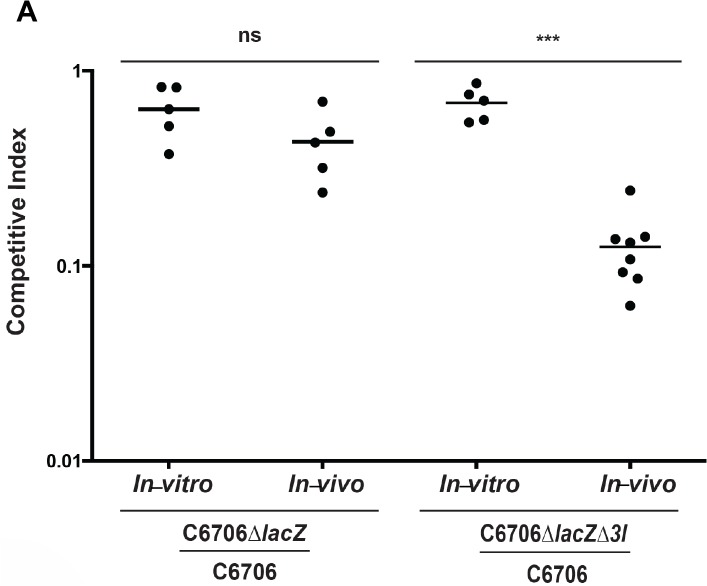
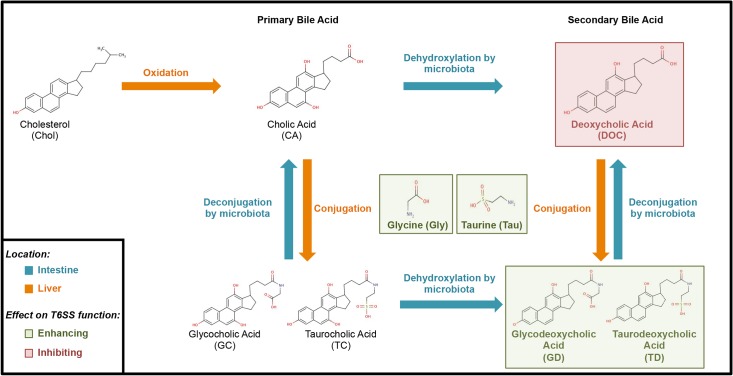
The causative agent of cholera, Vibrio cholerae, regulates its diverse virulence factors to thrive in the human small intestine and environmental reservoirs. Among this pathogen's arsenal of virulence factors is the tightly regulated type VI secretion system (T6SS). This system acts as an inverted bacteriophage to inject toxins into competing bacteria and eukaryotic phagocytes. V. cholerae strains responsible for the current 7th pandemic activate their T6SS within the host. We established that T6SS-mediated competition occurs upon T6SS activation in the infant mouse, and that this system is functional under anaerobic conditions. When investigating the intestinal host factors mucins (a glycoprotein component of mucus) and bile for potential regulatory roles in controlling the T6SS, we discovered that once mucins activate the T6SS, bile acids can further modulate T6SS activity. Microbiota modify bile acids to inhibit T6SS-mediated killing of commensal bacteria. This interplay is a novel interaction between commensal bacteria, host factors, and the V. cholerae T6SS, showing an active host role in infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Tacket CO, Losonsky G, Nataro JP, Cryz SJ, Edelman R, et al. (1993) Safety and immunogenicity of live oral cholera vaccine candidate CVD 110, a delta ctxA delta zot delta ace derivative of El Tor Ogawa Vibrio cholerae. J Infect Dis 168: 1536–1540. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
