

A MORN Repeat Protein Facilitates Protein Entry into the Flagellar Pocket of Trypanosoma brucei
- PMID: 26318396
- PMCID: PMC4621311
- DOI: 10.1128/EC.00094-15
A MORN Repeat Protein Facilitates Protein Entry into the Flagellar Pocket of Trypanosoma brucei
Abstract
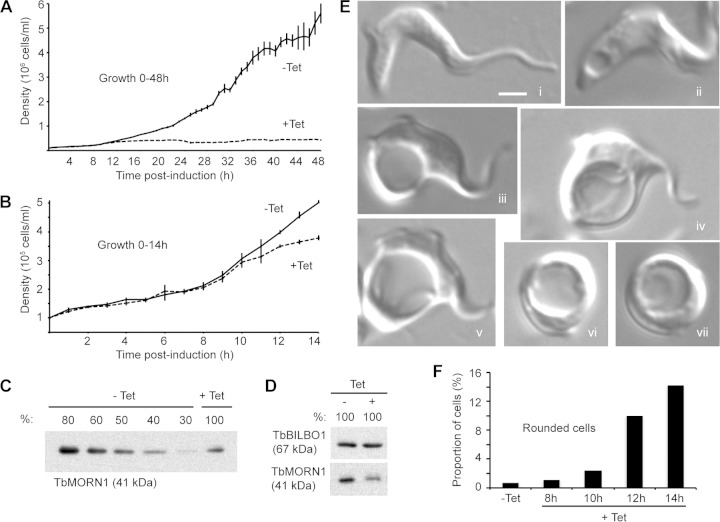
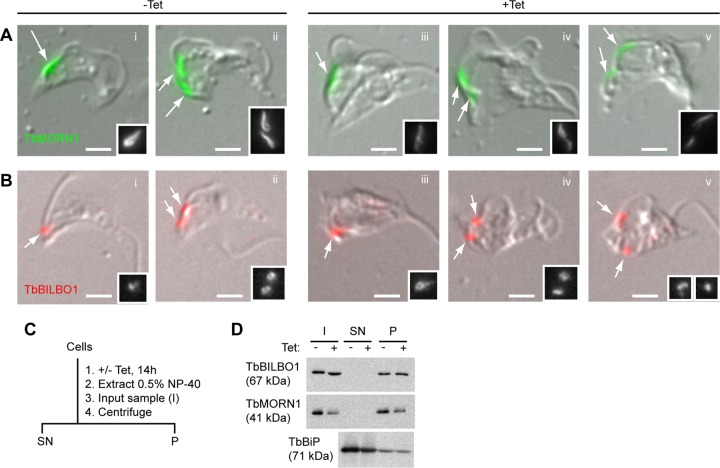
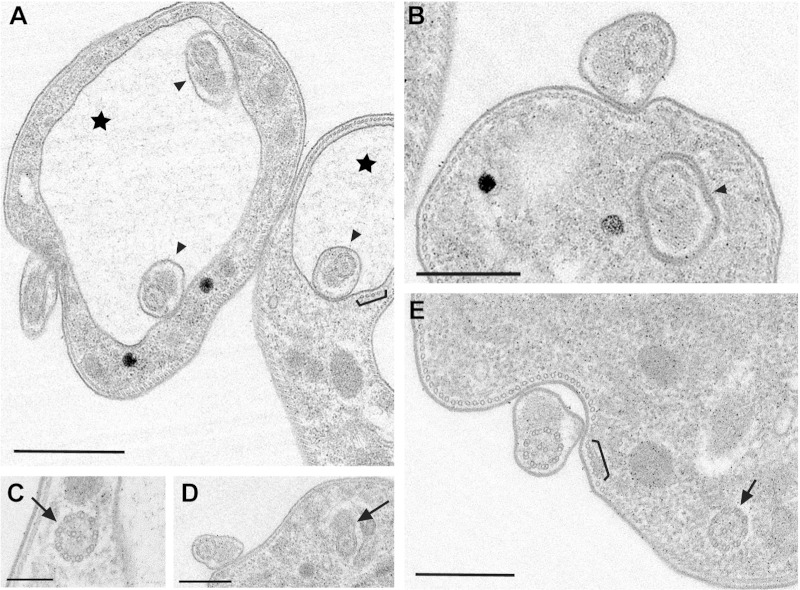
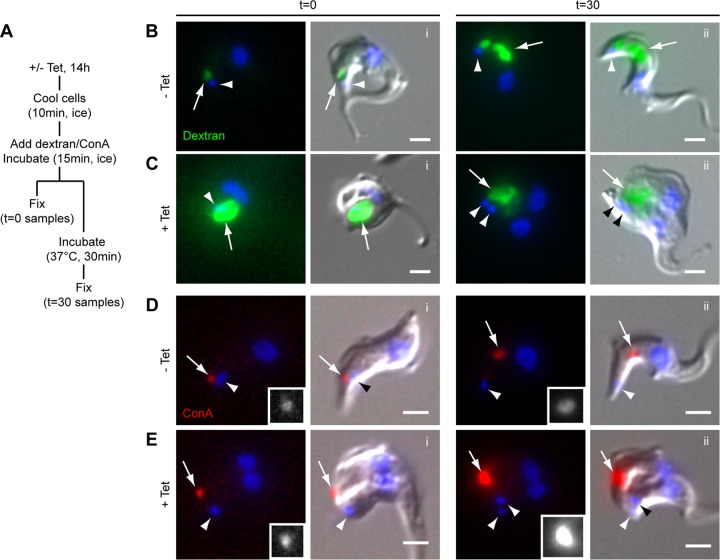
The parasite Trypanosoma brucei lives in the bloodstream of infected mammalian hosts, fully exposed to the adaptive immune system. It relies on a very high rate of endocytosis to clear bound antibodies from its cell surface. All endo- and exocytosis occurs at a single site on its plasma membrane, an intracellular invagination termed the flagellar pocket. Coiled around the neck of the flagellar pocket is a multiprotein complex containing the repeat motif protein T. brucei MORN1 (TbMORN1). In this study, the phenotypic effects of TbMORN1 depletion in the mammalian-infective form of T. brucei were analyzed. Depletion of TbMORN1 resulted in a rapid enlargement of the flagellar pocket. Dextran, a polysaccharide marker for fluid phase endocytosis, accumulated inside the enlarged flagellar pocket. Unexpectedly, however, the proteins concanavalin A and bovine serum albumin did not do so, and concanavalin A was instead found to concentrate outside it. This suggests that TbMORN1 may have a role in facilitating the entry of proteins into the flagellar pocket.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
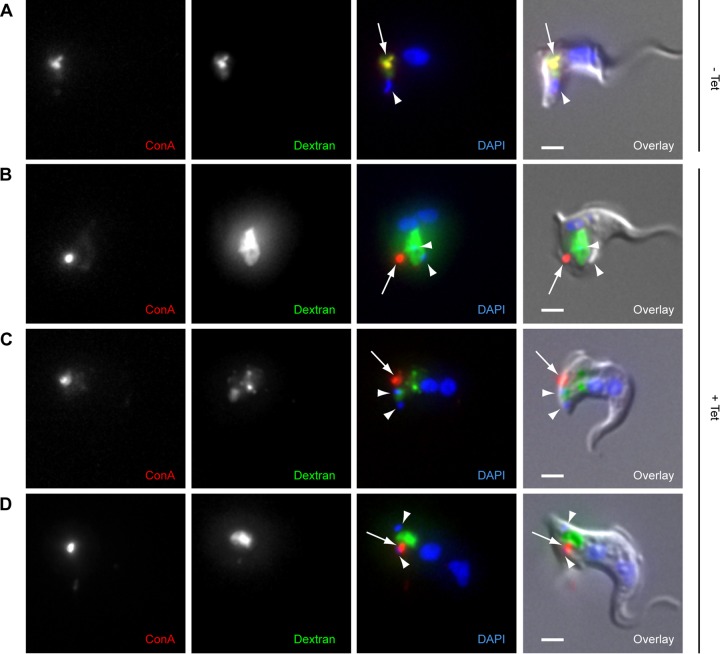
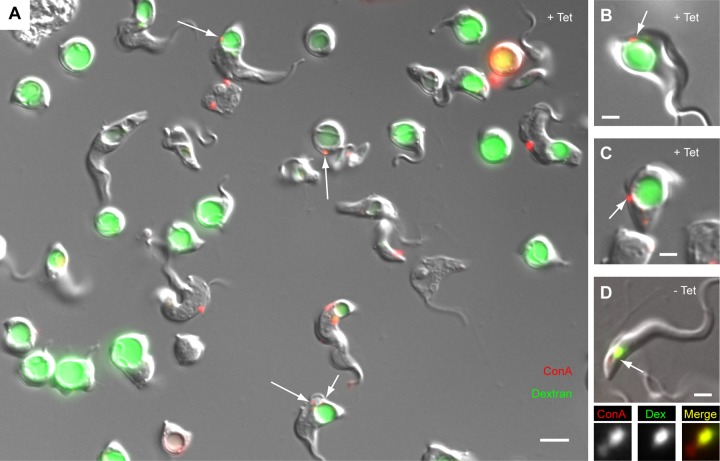
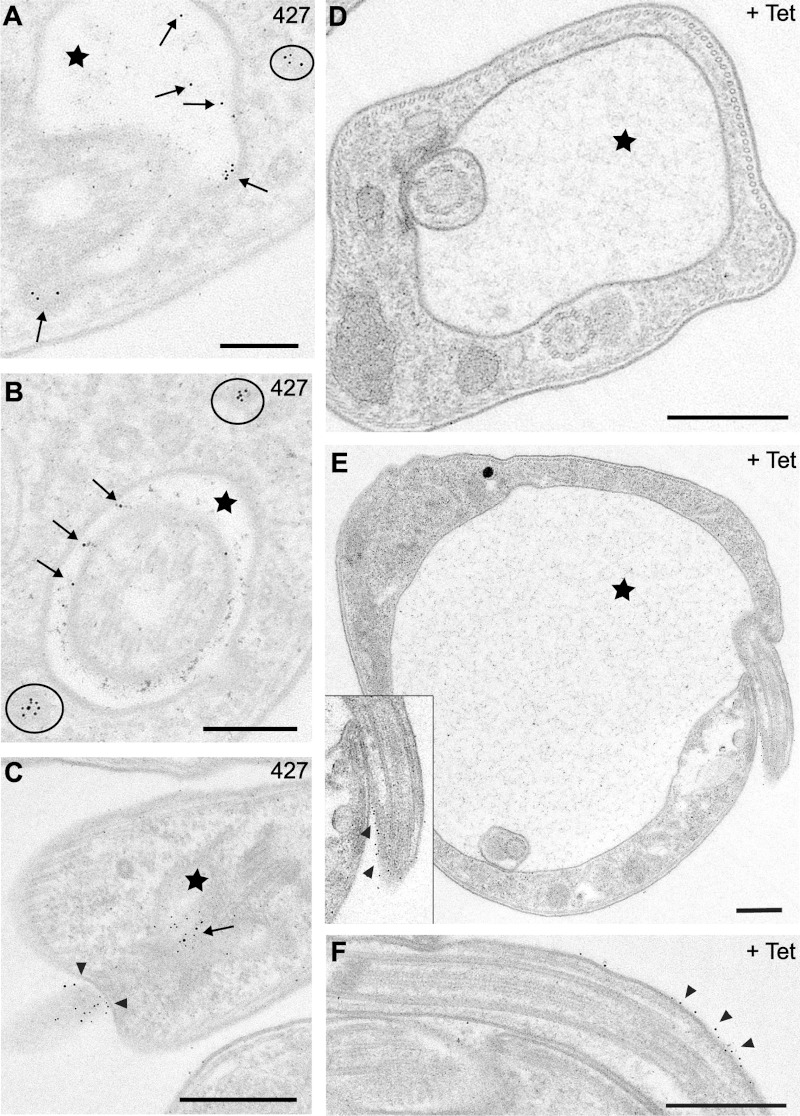
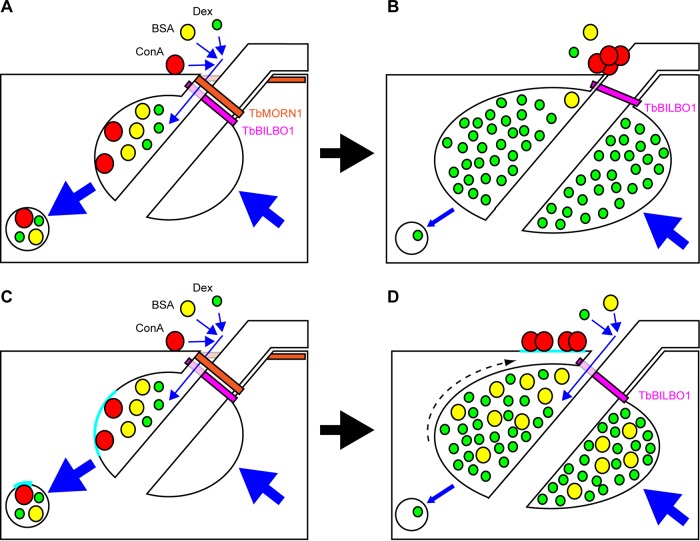
References
-
- Lacomble S, Vaughan S, Gadelha C, Morphew MK, Shaw MK, McIntosh JR, Gull K. 2009. Three-dimensional cellular architecture of the flagellar pocket and associated cytoskeleton in trypanosomes revealed by electron microscope tomography. J Cell Sci 122:1081–1090. doi: 10.1242/jcs.045740. - DOI - PMC - PubMed
-
- Grünfelder CG, Engstler M, Weise F, Schwarz H, Stierhof YD, Morgan GW, Field MC, Overath P. 2003. Endocytosis of a glycosylphosphatidylinositol-anchored protein via clathrin-coated vesicles, sorting by default in endosomes, and exocytosis via RAB11-positive carriers. Mol Biol Cell 14:2029–2040. doi: 10.1091/mbc.E02-10-0640. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
