

MicroRNA-27a decreases the level and efficiency of the LDL receptor and contributes to the dysregulation of cholesterol homeostasis
- PMID: 26318398
- PMCID: PMC4575910
- DOI: 10.1016/j.atherosclerosis.2015.08.023
MicroRNA-27a decreases the level and efficiency of the LDL receptor and contributes to the dysregulation of cholesterol homeostasis
Abstract
Rationale: A strong risk factor for atherosclerosis- the leading cause of heart attacks and strokes- is the elevation of low-density lipoprotein cholesterol (LDL-C) in blood. The LDL receptor (LDLR) is the primary pathway for LDL-C removal from circulation, and their levels are increased by statins -the main treatment for high blood LDL-C. However, statins have low efficiency because they also increase PCSK9 which targets LDLR for degradation. Since microRNAs have recently emerged as key regulators of cholesterol homeostasis, our aim was to identify potential microRNA-based therapeutics to decrease blood LDL-C and prevent atherosclerosis.
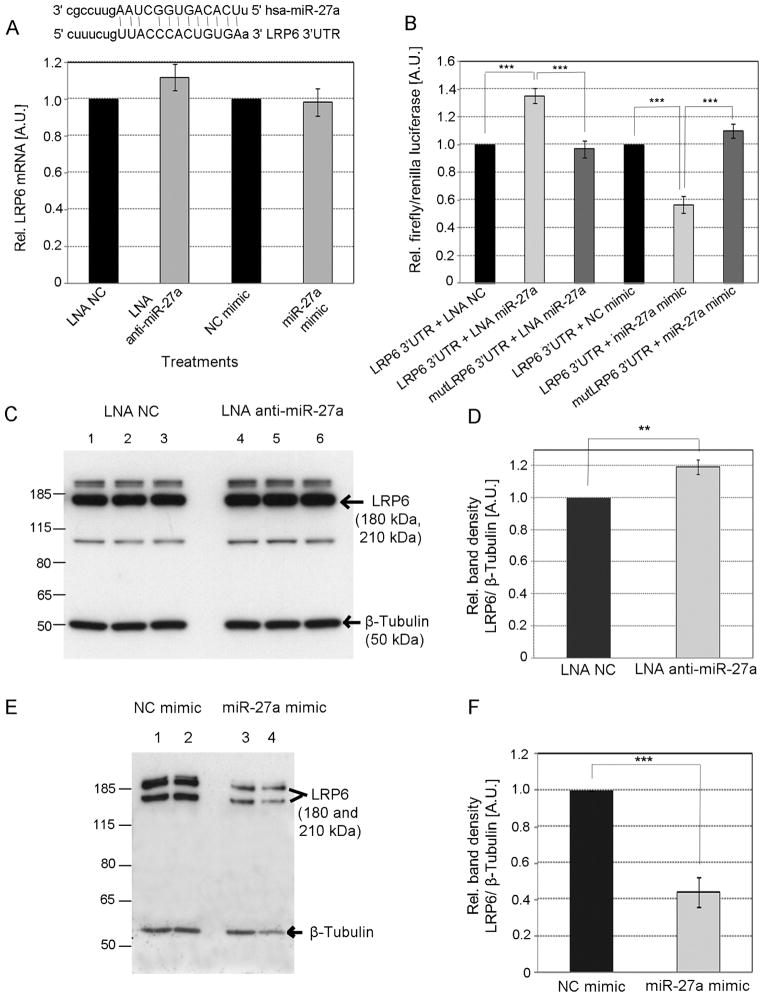
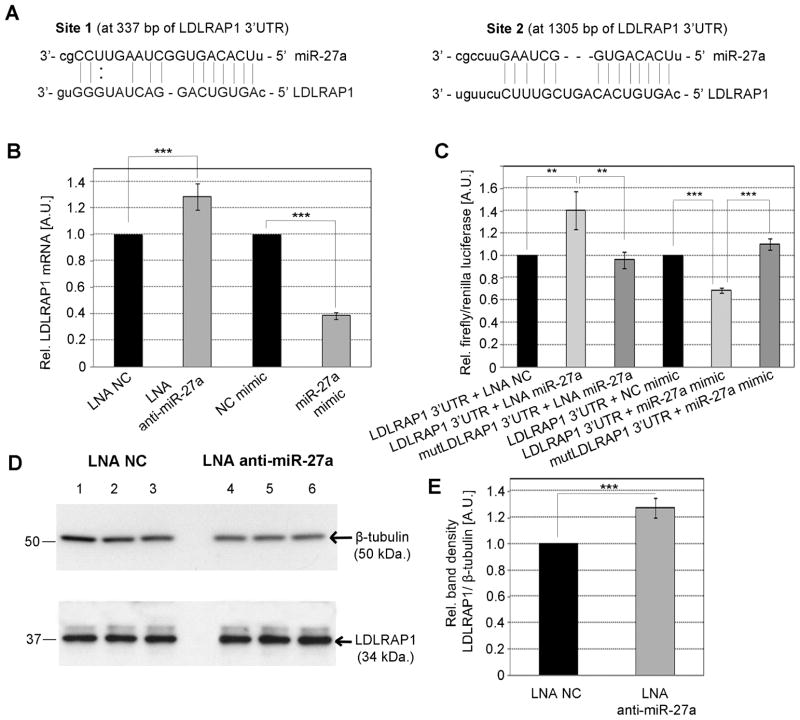
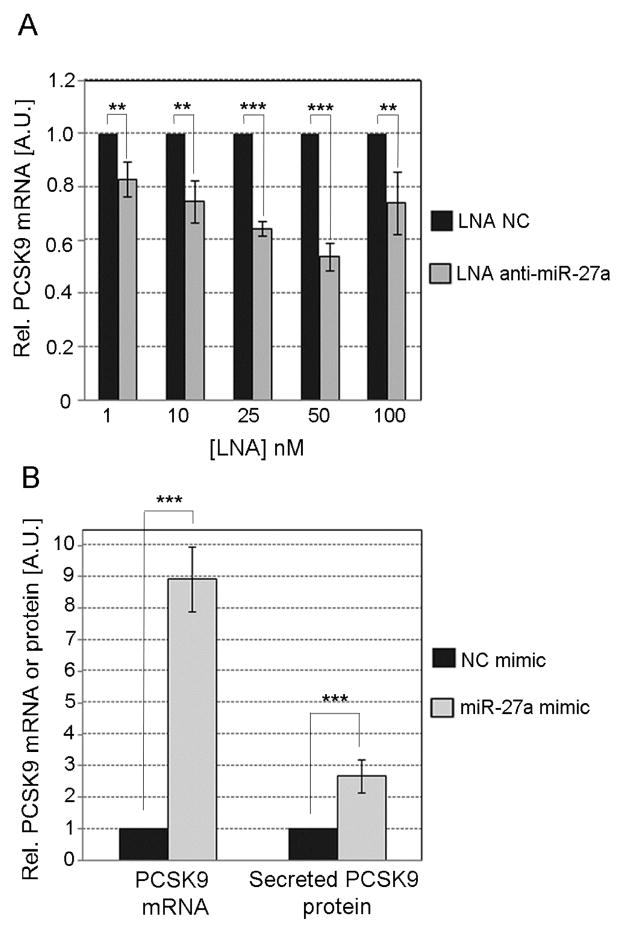
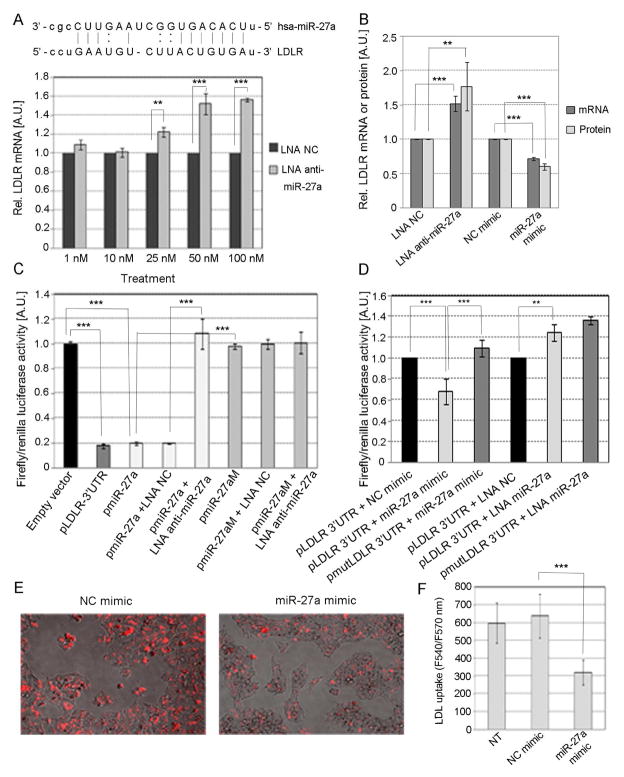
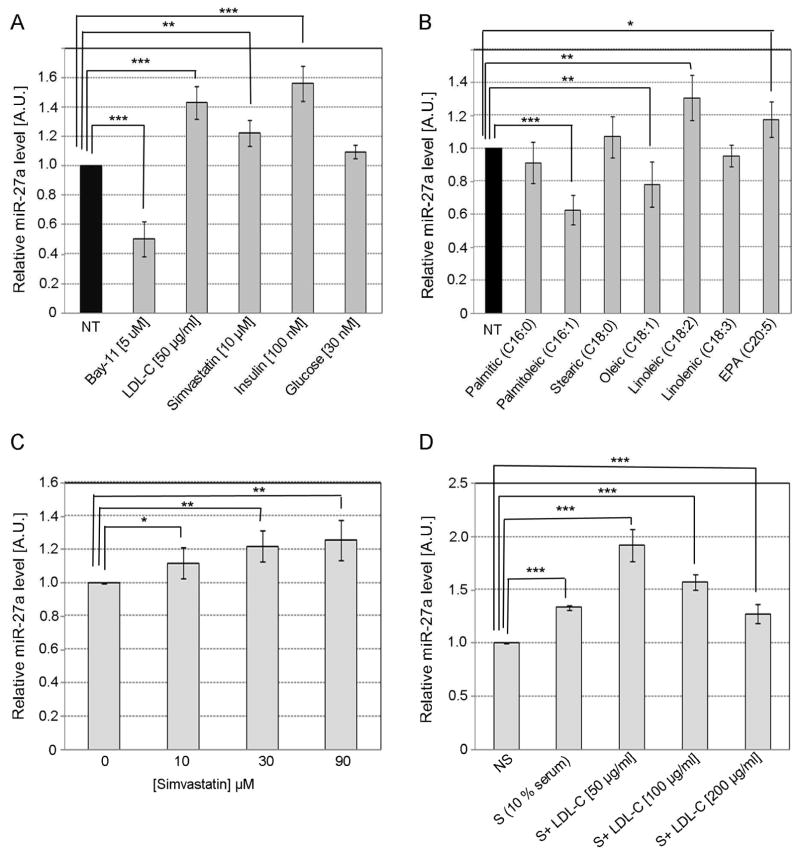
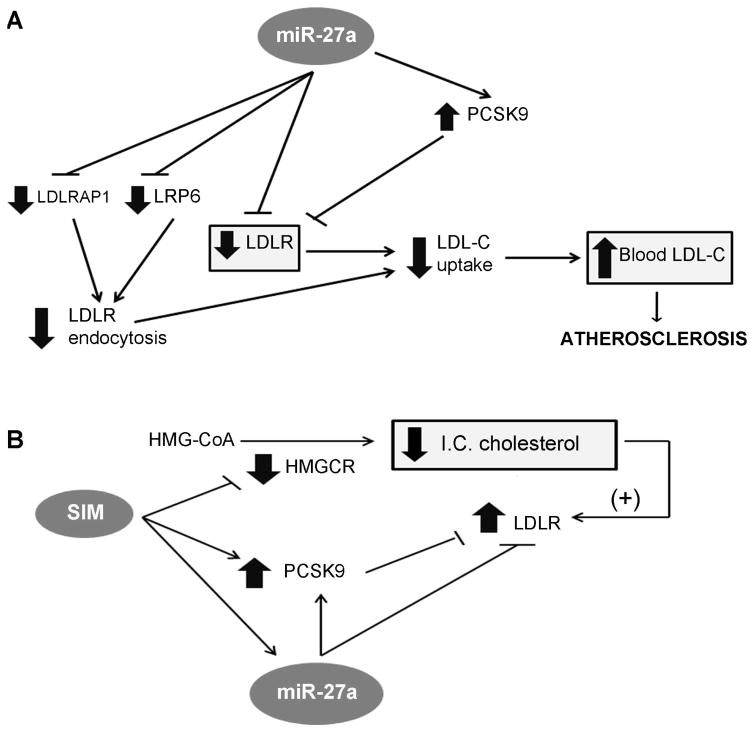
Methods and results: We over expressed and knocked down miR-27a in HepG2 cells to assess its effect on the expression of key players in the LDLR pathway using PCR Arrays, Elisas, and Western blots. We found that miR-27a decreases LDLR levels by 40% not only through a direct binding to its 3' untranslated region but also indirectly by inducing a 3-fold increase in PCSK9, which enhances LDLR degradation. Interestingly, miR-27a also directly decreases LRP6 and LDLRAP1, two other key players in the LDLR pathway that are required for efficient endocytosis of the LDLR-LDL-C complex in the liver. The inhibition of miR-27a using lock nucleic acids induced a 70% increase in LDLR levels and, therefore, it would be a more efficient treatment for hypercholesterolemia because of its desirable effects not only on LDLR but also on PCSK9.
Conclusion: The results presented here provide evidence supporting the potential of miR-27a as a novel therapeutic target for the prevention of atherosclerosis.
Keywords: Atherosclerosis; Cholesterol homeostasis; LDLR; LDLRAP1; LRP6; PCSK9; miR-27a.
Copyright © 2015 Elsevier Ireland Ltd. All rights reserved.
Figures
References
-
- Contu R, Latronico MV, Condorelli G. Circulating microRNAs as potential biomarkers of coronary artery disease: a promise to be fulfilled? Circ Res. 2010;107:573–574. - PubMed
-
- Mozaffarian D, Benjamin EJ, Go AS, et al. Heart disease and stroke statistics--2015 update: a report from the American Heart Association. Circulation. 2015;131:e29–322. - PubMed
-
- Abifadel M, Varret M, Rabes JP, et al. Mutations in PCSK9 cause autosomal dominant hypercholesterolemia. Nat Genet. 2003;34:154–156. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
