

Aluminum-Dependent Terminal Differentiation of the Arabidopsis Root Tip Is Mediated through an ATR-, ALT2-, and SOG1-Regulated Transcriptional Response
- PMID: 26320227
- PMCID: PMC4815104
- DOI: 10.1105/tpc.15.00172
Aluminum-Dependent Terminal Differentiation of the Arabidopsis Root Tip Is Mediated through an ATR-, ALT2-, and SOG1-Regulated Transcriptional Response
Abstract
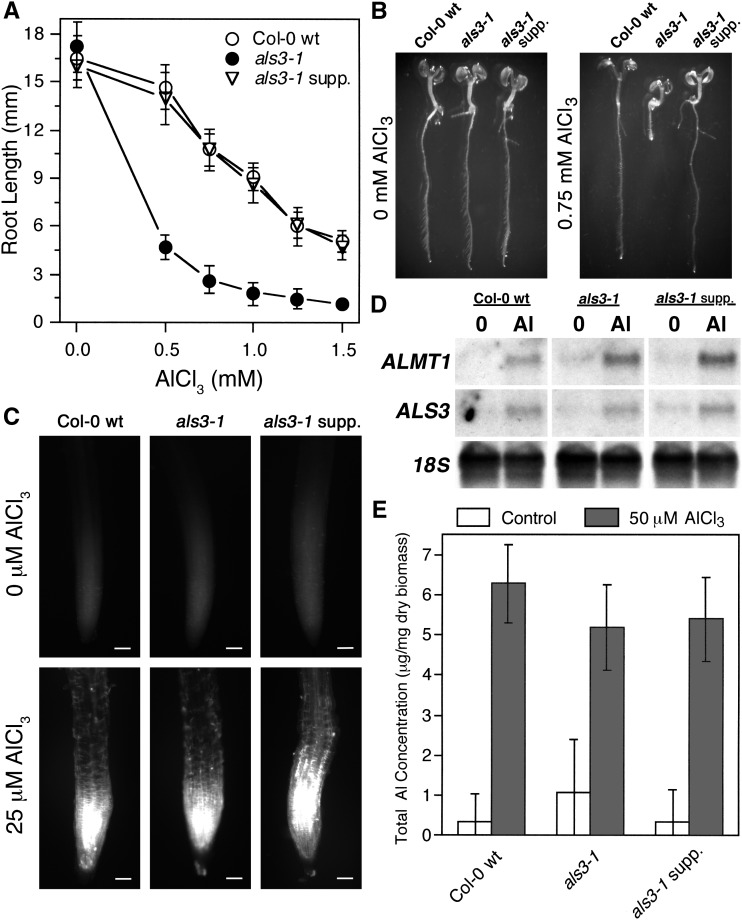
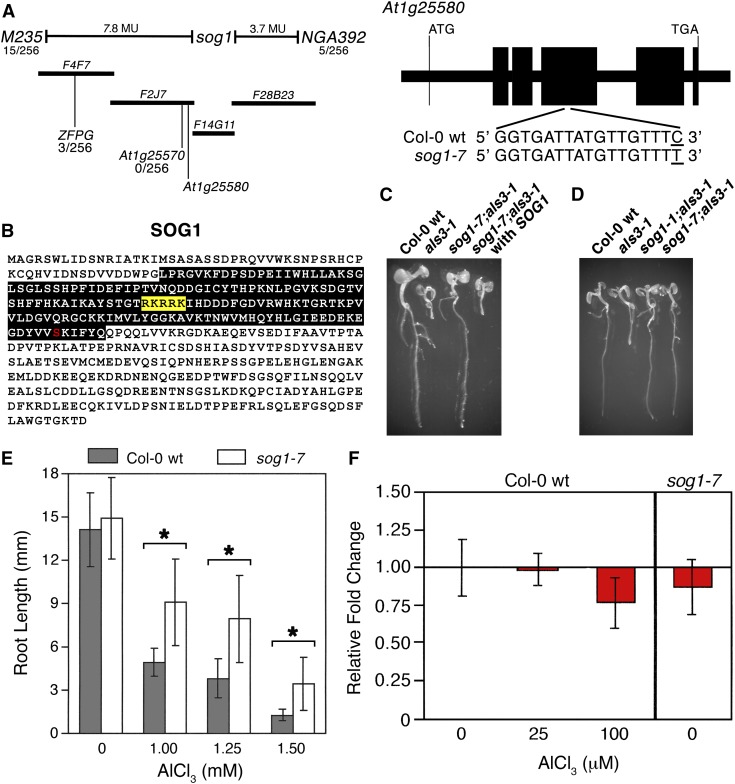
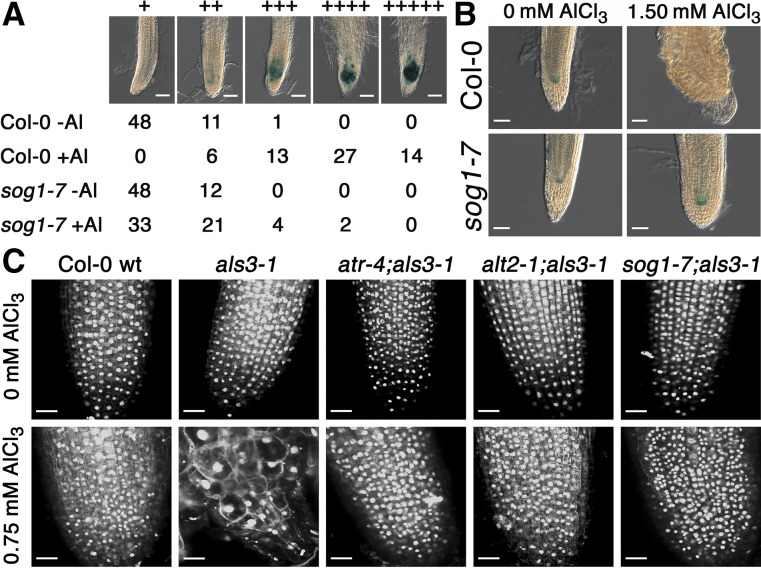
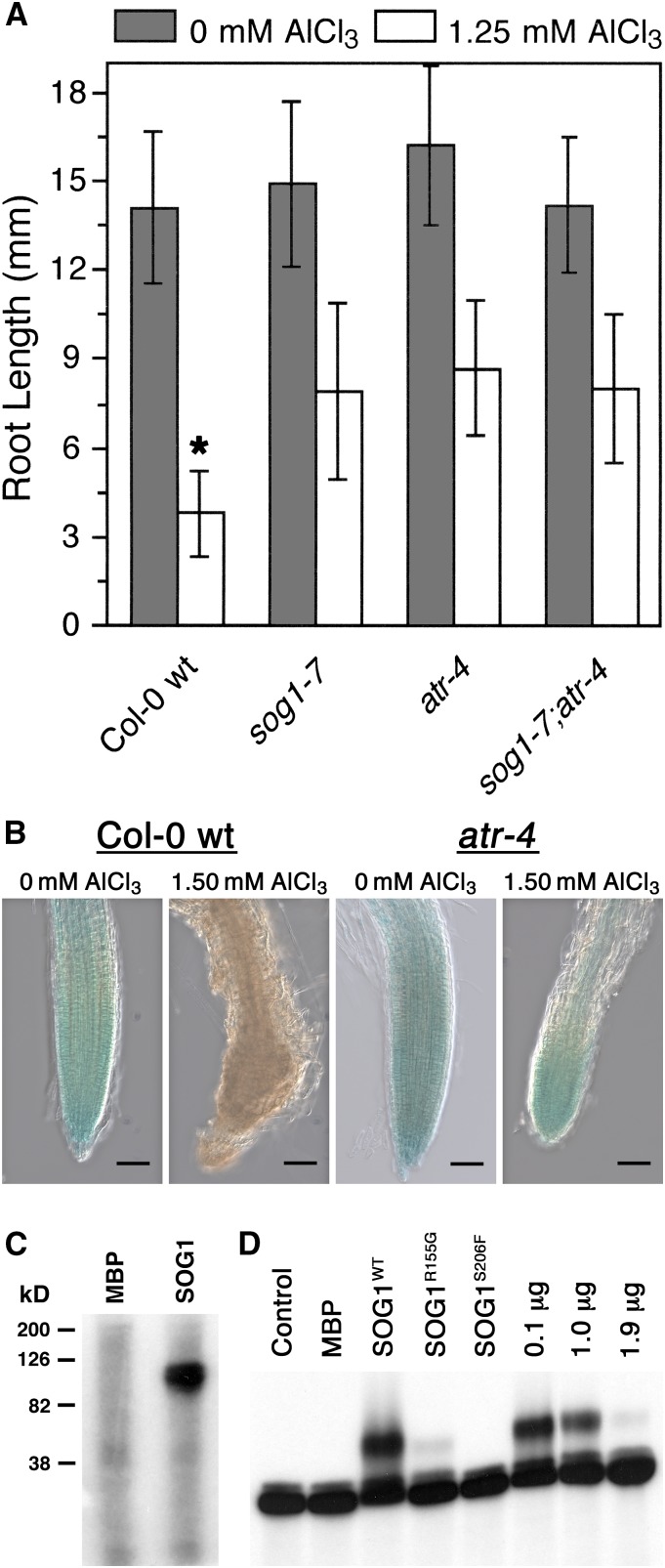
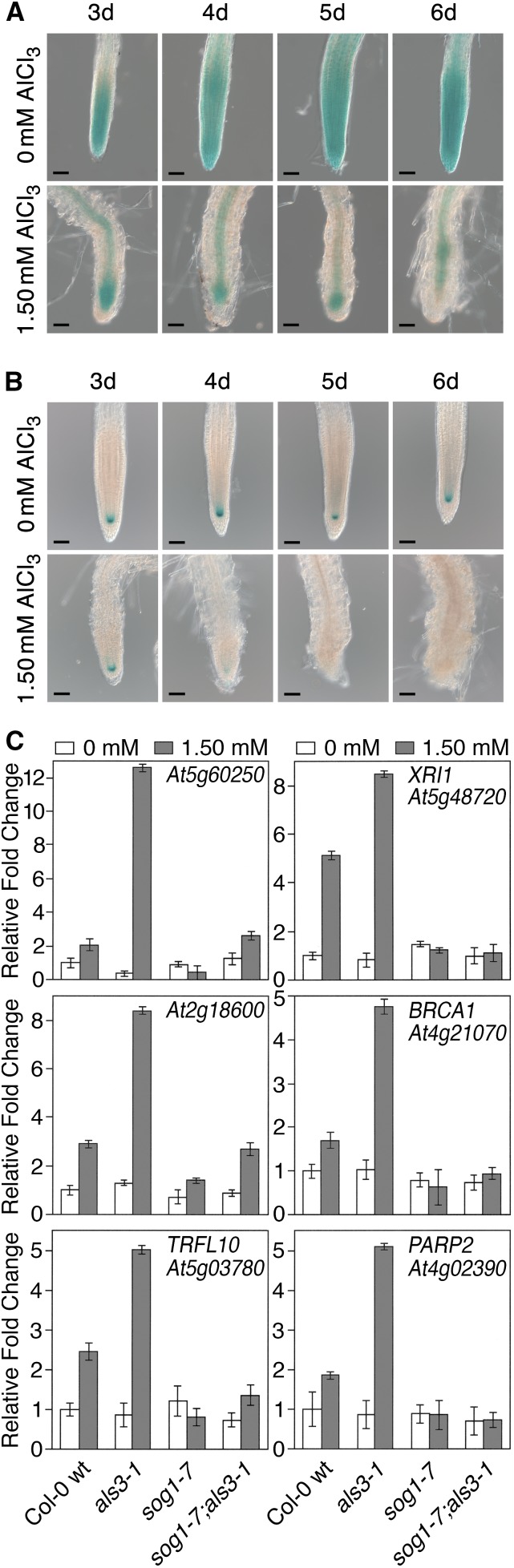
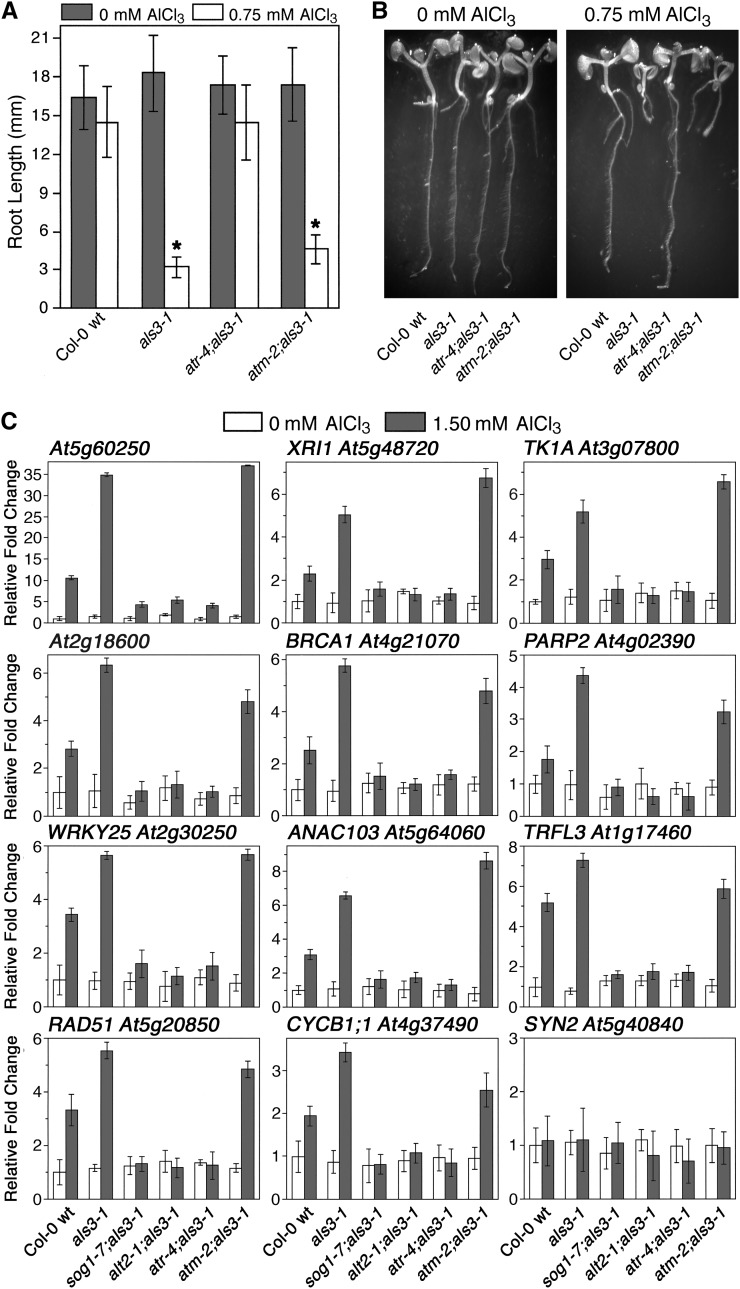
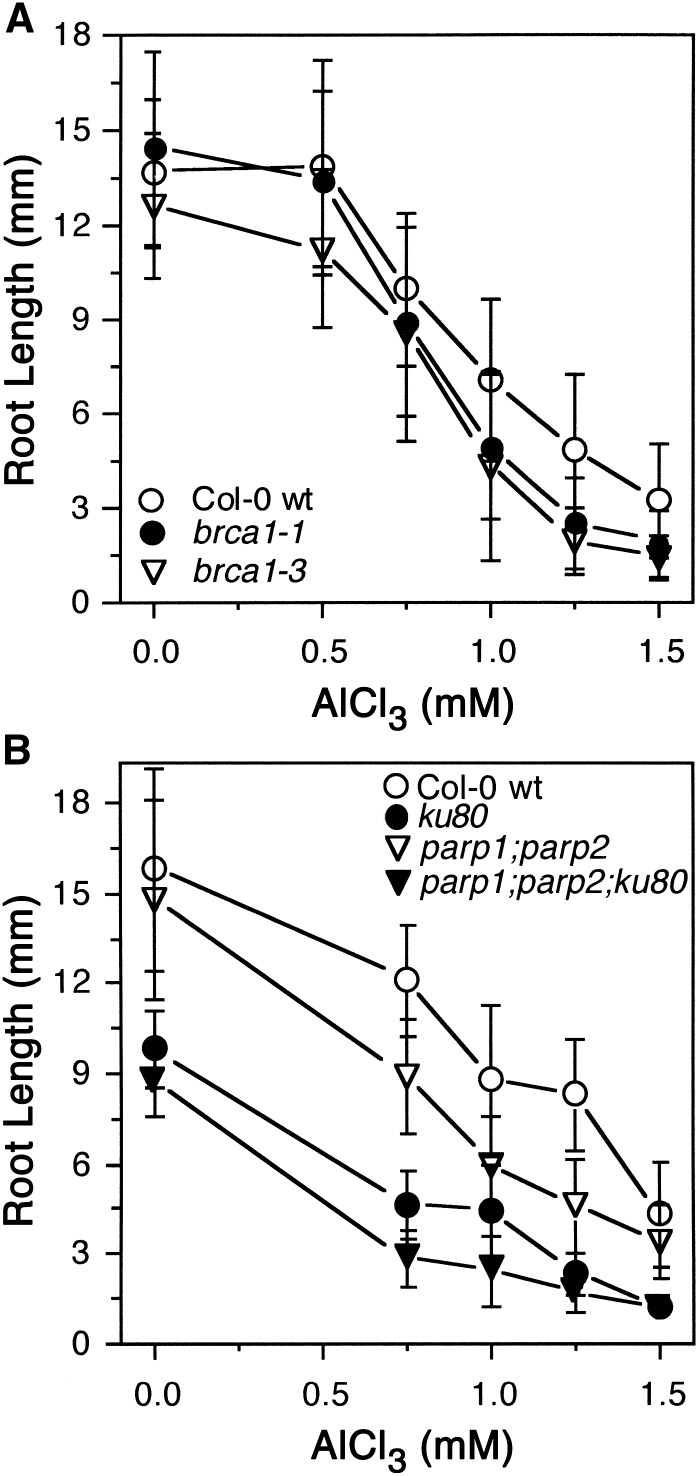
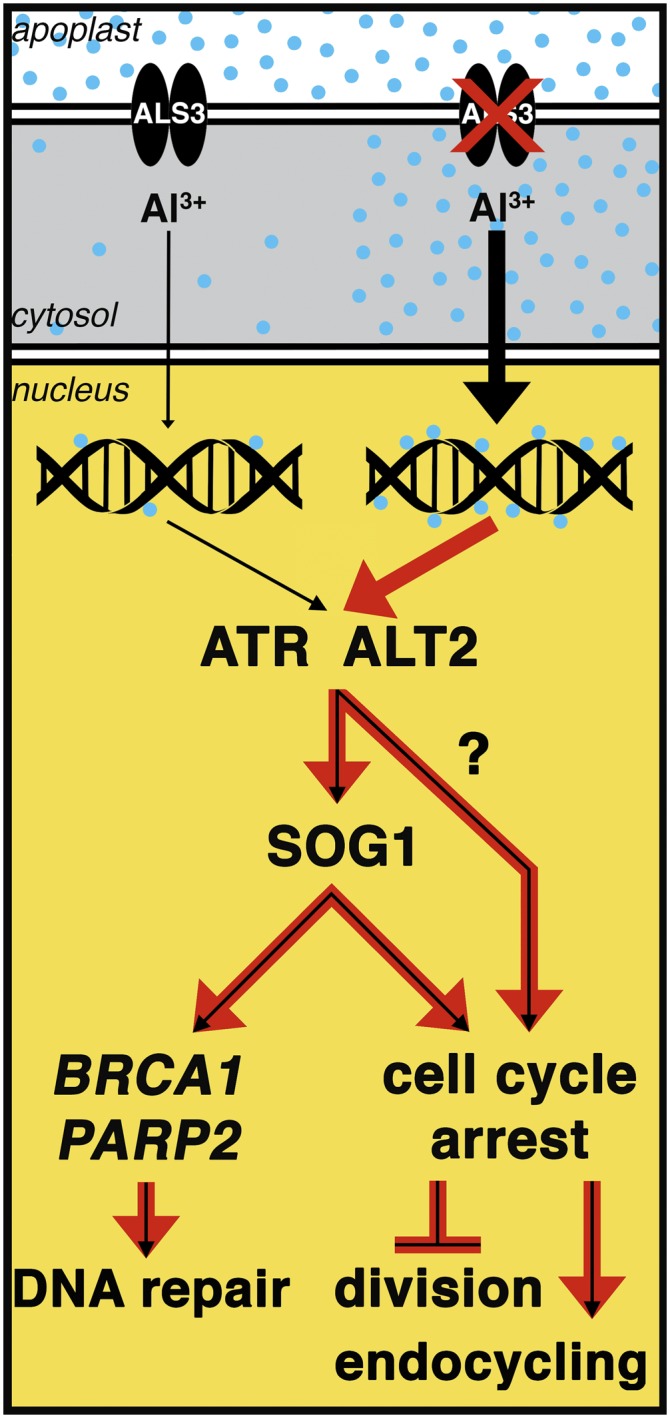
By screening for suppressors of the aluminum (Al) hypersensitive Arabidopsis thaliana mutant als3-1, it was found that mutational loss of the Arabidopsis DNA damage response transcription factor SUPPRESSOR OF GAMMA RESPONSE1 (SOG1) confers increased Al tolerance similar to the loss-of-function mutants for the cell cycle checkpoint genes ATAXIA TELANGIECTASIA AND RAD3 RELATED (ATR) and ALUMINUM TOLERANT2 (ALT2). This suggests that Al-dependent terminal differentiation of the root tip is an active process resulting from activation of the DNA damage checkpoint by an ATR-regulated pathway, which functions at least in part through SOG1. Consistent with this, ATR can phosphorylate SOG1 in vitro. Analysis of SOG1's role in Al-dependent root growth inhibition shows that sog1-7 prevents Al-dependent quiescent center differentiation and endoreduplication in the primary root tip. Following Al exposure, SOG1 increases expression of several genes previously associated with DNA damage, including BRCA1 and PARP2, with gel-shift analysis showing that SOG1 can physically associate with the BRCA1 promoter in vitro. Al-responsive expression of these SOG1-regulated genes requires ATR and ALT2, but not ATAXIA TELANGIECTASIA MUTATED, thus demonstrating that in response to chronic Al exposure, ATR, ALT2, and SOG1 function together to halt root growth and promote terminal differentiation at least in part in a transcription-dependent manner.
© 2015 American Society of Plant Biologists. All rights reserved.
Figures
Similar articles
-
SUV2, which encodes an ATR-related cell cycle checkpoint and putative plant ATRIP, is required for aluminium-dependent root growth inhibition in Arabidopsis.Plant Cell Environ. 2017 Sep;40(9):1849-1860. doi: 10.1111/pce.12992. Epub 2017 Jul 12. Plant Cell Environ. 2017. PMID: 28556304
-
The Arabidopsis cell cycle checkpoint regulators TANMEI/ALT2 and ATR mediate the active process of aluminum-dependent root growth inhibition.Plant Cell. 2012 Feb;24(2):608-21. doi: 10.1105/tpc.112.095596. Epub 2012 Feb 17. Plant Cell. 2012. PMID: 22345493 Free PMC article.
-
An insight into the mechanism of DNA damage response in plants- role of SUPPRESSOR OF GAMMA RESPONSE 1: An overview.Mutat Res. 2020 Jan-Apr;819-820:111689. doi: 10.1016/j.mrfmmm.2020.111689. Epub 2020 Jan 23. Mutat Res. 2020. PMID: 32004947 Review.
-
The stem cell niche transcription factor ETHYLENE RESPONSE FACTOR 115 participates in aluminum-induced terminal differentiation in Arabidopsis roots.Plant Cell Environ. 2024 Nov;47(11):4432-4448. doi: 10.1111/pce.15032. Epub 2024 Jul 15. Plant Cell Environ. 2024. PMID: 39007549
-
SOG1: a master regulator of the DNA damage response in plants.Genes Genet Syst. 2016;90(4):209-16. doi: 10.1266/ggs.15-00011. Epub 2015 Nov 26. Genes Genet Syst. 2016. PMID: 26617076 Review.
Cited by
-
Different effects of gellan gum and agar on change in root elongation in Arabidopsis thaliana by polyploidization: the key role of aluminum.J Plant Res. 2023 Mar;136(2):253-263. doi: 10.1007/s10265-023-01435-x. Epub 2023 Jan 23. J Plant Res. 2023. PMID: 36689102
-
Genome-wide identification and expression analysis of the NAC transcription factor family in tomato (Solanum lycopersicum) during aluminum stress.BMC Genomics. 2020 Apr 7;21(1):288. doi: 10.1186/s12864-020-6689-7. BMC Genomics. 2020. PMID: 32264854 Free PMC article.
-
The Plant DNA Damage Response: Signaling Pathways Leading to Growth Inhibition and Putative Role in Response to Stress Conditions.Front Plant Sci. 2019 May 17;10:653. doi: 10.3389/fpls.2019.00653. eCollection 2019. Front Plant Sci. 2019. PMID: 31164899 Free PMC article. Review.
-
The ATR-WEE1 kinase module promotes SUPPRESSOR OF GAMMA RESPONSE 1 translation to activate replication stress responses.Plant Cell. 2023 Aug 2;35(8):3021-3034. doi: 10.1093/plcell/koad126. Plant Cell. 2023. PMID: 37159556 Free PMC article.
-
Increased Phosphorylation of Ser-Gln Sites on SUPPRESSOR OF GAMMA RESPONSE1 Strengthens the DNA Damage Response in Arabidopsis thaliana.Plant Cell. 2017 Dec;29(12):3255-3268. doi: 10.1105/tpc.17.00267. Epub 2017 Dec 5. Plant Cell. 2017. PMID: 29208704 Free PMC article.
References
-
- Chandran D., Sharopova N., Ivashuta S., Gantt J.S., Vandenbosch K.A., Samac D.A. (2008). Transcriptome profiling identified novel genes associated with aluminum toxicity, resistance and tolerance in Medicago truncatula. Planta 228: 151–166. - PubMed
-
- Colón-Carmona A., You R., Haimovitch-Gal T., Doerner P. (1999). Technical advance: spatio-temporal analysis of mitotic activity with a labile cyclin-GUS fusion protein. Plant J. 20: 503–508. - PubMed
-
- Culligan K.M., Robertson C.E., Foreman J., Doerner P., Britt A.B. (2006). ATR and ATM play both distinct and additive roles in response to ionizing radiation. Plant J. 48: 947–961. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
