

Comment
doi: 10.1093/jhered/esv056.
Epub 2015 Aug 29.
Meiotic Models to Explain Classical Linkage, Pseudolinkage, and Chromosomal Pairing in Tetraploid Derivative Salmonid Genomes: II. Wright is Still Right
Affiliations
- PMID: 26320244
- PMCID: PMC4642675
- DOI: 10.1093/jhered/esv056
Item in Clipboard
Comment
Meiotic Models to Explain Classical Linkage, Pseudolinkage, and Chromosomal Pairing in Tetraploid Derivative Salmonid Genomes: II. Wright is Still Right
J Hered.
2015 Nov-Dec.
No abstract available
Figures
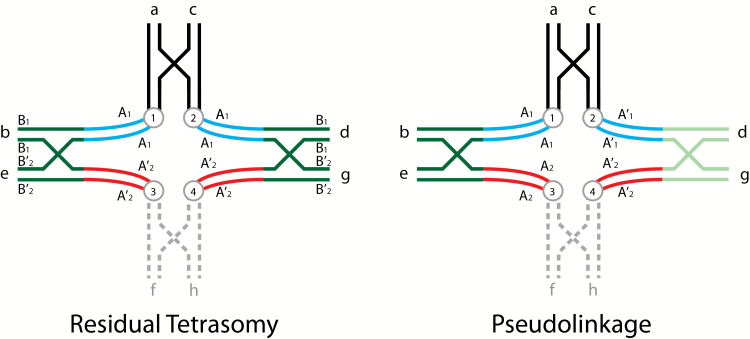
Diagrammatic illustrations of tetravalent pairing of homologous regions of homeologous chromosome arms illustrating both residual tetrasomy and pseudolinkage in salmonid males (adapted from Wright et al. 1983). Residual tetrasomy is illustrated on the left side with chromosomal arms labeled in lower case letters and color coded, centromeres appear as circles with letters, 2 loci are shown, where the A locus is proximal to the centromere (centromeric) and the B locus is distal (telomeric), and a single crossover event is shown in the telomeric region of each pair of arms. Arms f and h are shown in broken grey lines to allow the reader to view this tetravalent as 2 metacentrics (ab and cd) and either 2 acrocentrics (e and g) or another set of metacentrics (ef and gh). The a and c arms, the b and d arms, and the e and g are homologous (as would be f and h). Arms b and d are homeologous with respect to e and g, that is, they were ancestrally homologous, but have diverged. Using arm b as an example, it is composed of 2 general regions where the dark green telomeric region is homologous to not only the telomeric region of d, but it is also still homologous with the telomeric regions of e and g (also dark green), and can pair homologously in the telomeric region (under the male meiotic environment) with either e or g. The centromeric area of b (blue) is homologous with the centromeric region of d and they can pair as bivalents. The centromeric area of b is homeologous with e and g (red) and cannot pair in these regions with e and g. The homologous telomeric regions (dark green) are residually tetrasomic and the centromeric regions (blue and red) are disomic, that is, the A loci have diverged and are respectively disomic and the B loci can exhibit disomic or tetrasomic pairing and are isoloci (sharing the same alleles). We show these centromeric areas separated from each other in this figure. When these arms separate via alternate disjunction centromeres 1 and 4 will go to one meiotic pole and centromeres 2 and 3 will go to the other pole. If this is one set of metacentrics and one set of acrocentrics the chiasma will terminalize forming a long rod, or a ring if there are 2 sets of metacentrics involved. In some meioses this tetravalent will form and in other meioses only the 2 respective bivalents will form. The ratio of bivalent versus tetravalent formation will give the intermediate results for the B locus shown for males in Table 1 of Allendorf et al. (2015). The 2 A loci will show random assortment. The pseudolinkage figure is similar to that shown for residual tetrasomy, except that the A locus alleles are labeled differently where the A comes from one parent and the A’ from the other parent, the B locus is dropped, and homologous distal regions of the homeologous arms are shaded differently. For discussion purposes, we will consider this figure to represent meiotic pairing in a male splake (hybrid between lake and brook/speckled trout). Note: the parental genomes could have come from divergent populations of the same species. Arms c, d, g(gh) come from the lake trout parent and arms a, b, e(ef) from the brook trout parent. In this case a and c, both shown in black will pair since they have no homeologues, (similarly f and h), even though there may be substantial differentiation. The centromeric regions of b and d are both shown in blue and likewise the centromeric regions of e and g are still both shown in red, even they have clearly diverged from one another, in order to reduce the number of colors. The telomeric region of arm b (dark green) will preferentially pair with e (dark green), while arm d (light green) will pair with g (light green) because of their greater homology. This preferential pairing is what makes pseudolinkage a specialized case of residual tetrasomy. Alternate disjunction (centromeres 1 and 4 going to one pole and centromeres 2 and 3 going to the other) will lead to the formation of only non-parental gametes A’1 and A2 and A1 and A’2 every time this tetravalent forms since there is no crossing over between the A locus and the centromere. Parental and non-parental gametes of the 2 unlinked A loci will be produced in equal frequencies whenever bivalents of these chromosome are formed. Again, the ratio of formation of bivalent versus tetravalent pairing will determine the extent of the pseudolinkage of the 2 disomic A loci. We (and Allendorf et al. 2015) have limited allelic variation within these models to illustrate the phenomena without introducing the complexity of within locus variability. The reader is encouraged to add allelic variation at each locus within and between each parental chromosome to see the gametic possibilities.
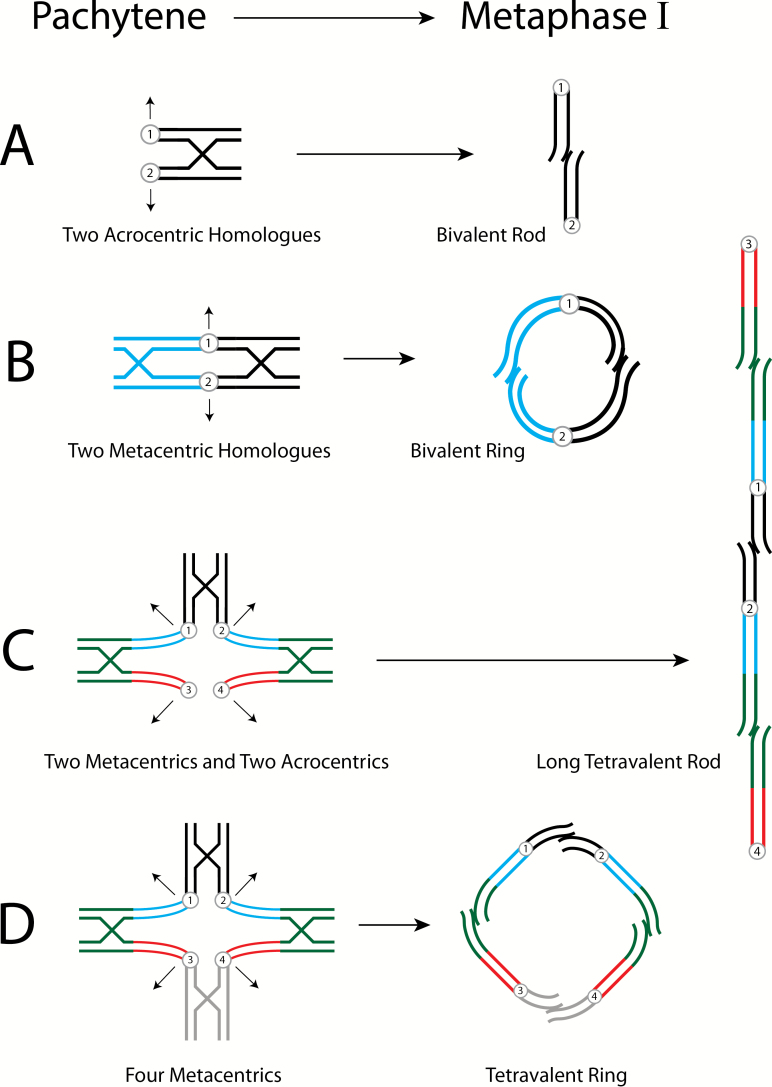
Diagrammatic interpretation of male salmonid pairing configurations, Pachytene to Metaphase of Meiosis I. Sister chromatids are adjacent, circles with numbers denote centromeres. Recombination and chiasmata terminalization indicated by cross structures; arrows indicate direction of the chromosomes ‘opening’ to opposite poles. (A) Bivalent pairing of 2 acrocentrics, disomic homologues (black) opens to a small rod. (B) Bivalent pairing of 2 metacentrics, disomic homologues (blue and black) opens to a small ring. (C) Tetravalent pairing involving 2 metacentrics (homologues) and 2 acrocentrics (homologues) which opens to a long mulitvalent rod. The homologous telomeric ends of the homeologous metacentrics and acrocentrics pair, while the disomic centromeric regions do not. See Figure 1 for a more detailed description of the tetravalent. It is important to remember in all of these figures that they are 2-dimensional compressions of 3-dimensional phenomena. Centromeres 1 and 4 which are moving to one meiotic pole can be considered to be above the page and centromeres 2 and 3 which are moving to the opposite pole can be considered to be below the page. (D) Tetravalent pairing of 4 metacentrics that opens to a large tetravalent ring. This is similar to 2C except the acrocentric pair is replaced by a pair of metacentrics.
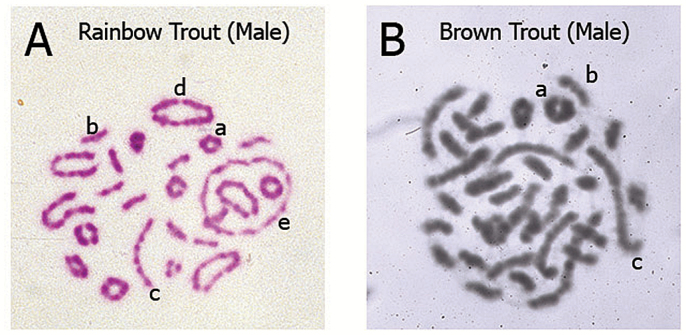
Meiosis I metaphase stage chromosome pairing configurations of male salmonids with different karyotype architecture: bivalent and multivalent pairing results from homologous and homeologous ancestral relationships. Explanation of the configurations is provided in the context of major events of Meiosis I. Chromosome regions that synapse (pair) and recombine (undergo crossing-over) during Prophase I (pachytene) will move to align and orient on the Metaphase I equitorial plate via the action of centriole-derived spindle fibers which attach to the centromeres and create tension on the chromosomes. The polymerization/depolymerization action of the spindle fibers results in the pulling of alternating chromosomes toward the opposite poles of the dividing cell. Thus, a cross structure in Prophase I involving 4 metacentrics becomes a large ring of 4 chromosomes (tetravalent) as the chiasmata terminalize (move to the ends of the chromosome), see Figure 2D. Ultimately the maternal and paternal chromosomes segregate (assort) independently and accurately during the reductional division of Meiosis I (one diploid 2n cell divides to produce 2 haploid 1n cells). The types of pairing configurations observed relates directly to the architecture of the karyotype. That is, the number of metacentrics and acrocentrics found in the karyotype which varies among salmonid species due to the variable number of Robertsonian centric fusions which occurred during species radiation following the ancestral tetraploid event. (A) Representative example of rainbow trout (Oncorhynchus mykiss) bivalent and multivalent pairing configurations (Giemsa stained, color image). Typically, rainbow trout show several large multivalent rings (4, 6, or 8 metacentrics), a lesser number of long multivalent rods (2 metacentrics/2 acrocentrics) and bivalent pairing (small rings of 2 metacentrics and small rods of 2 acrocentrics). This image is of a cell from a fish from a Benner Springs Hatchery (PA) population, 2n = 64 (40 metacentrics, 24 acrocentrics; 104 arms). (B) Representative example of brown trout (Salmo trutta) bivalent and multivalent pairing configurations (Giemsa stained, black/white image). Typically, brown trout show several long multivalent rods (2 metacentrics/2 acrocentrics) and bivalent pairing (a low number of small rings of 2 metacentrics and numerous small rods of 2 acrocentrics). This image is of a cell from a fish from a Benner Springs Hatchery (PA) population, 2n = 80 (20 metacentrics, 60 acrocentrics; 100 arms). Representative configurations indicated by the lowercase letters in A and B: (a) small ring bivalent of 2 metacentrics pairing (b) small rod bivalent of 2 acrocentrics pairing (c) long rod multivalent (tetravalent) of an acrocentric–metacentric–metacentric–acrocentric pairing, common in species with high numbers of acrocentrics (d) large ring multivalent (tetravalent) of 4 metacentrics pairing, common in species with high numbers of metacentrics (e) large ring multivalent (hexavalent) of 6 metacentrics pairing, common in species with high numbers of metacentrics. The chromosome preparations shown were created from cells harvested from male salmonids in 1980 (MED) in the cytogenetics laboratory of the late James E. Wright, Jr., at The Pennsylvania State University, State College, PA and supported by an NSF grant to JEW in 1979.
Comment in
-
Response to May and Delany: We Never Said Wright was Wrong.J Hered. 2015 Nov-Dec;106(6):767-8. doi: 10.1093/jhered/esv072. Epub 2015 Aug 29. J Hered. 2015. PMID: 26320245 No abstract available.
Comment on
-
Effects of crossovers between homeologs on inheritance and population genomics in polyploid-derived salmonid fishes.J Hered. 2015 May-Jun;106(3):217-27. doi: 10.1093/jhered/esv015. Epub 2015 Apr 2. J Hered. 2015. PMID: 25838153 Review.
References
-
- Allendorf FW, Bassham S, Cresko WA, Limborg MT, Seeb LW, Seeb JE. 2015. Effects of crossovers between homeologs on inheritance and population genomics in polyploid-derived salmonid fishes. J Hered. 106:217–227. - PubMed
-
- Allendorf FW, Thorgaard G. 1984. Polyploidy and the evolution of salmonid fishes. In: Turner BJ, editor. The evolutionary genetics of fishes. New York: (NY: ): Plenum Press; p. 1–53.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
