

Inhibitory and excitatory axon terminals share a common nano-architecture of their Cav2.1 (P/Q-type) Ca(2+) channels
- PMID: 26321916
- PMCID: PMC4531237
- DOI: 10.3389/fncel.2015.00315
Inhibitory and excitatory axon terminals share a common nano-architecture of their Cav2.1 (P/Q-type) Ca(2+) channels
Abstract
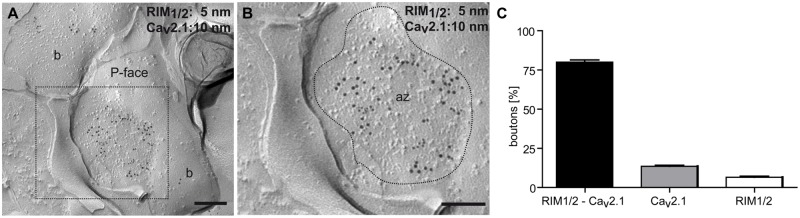
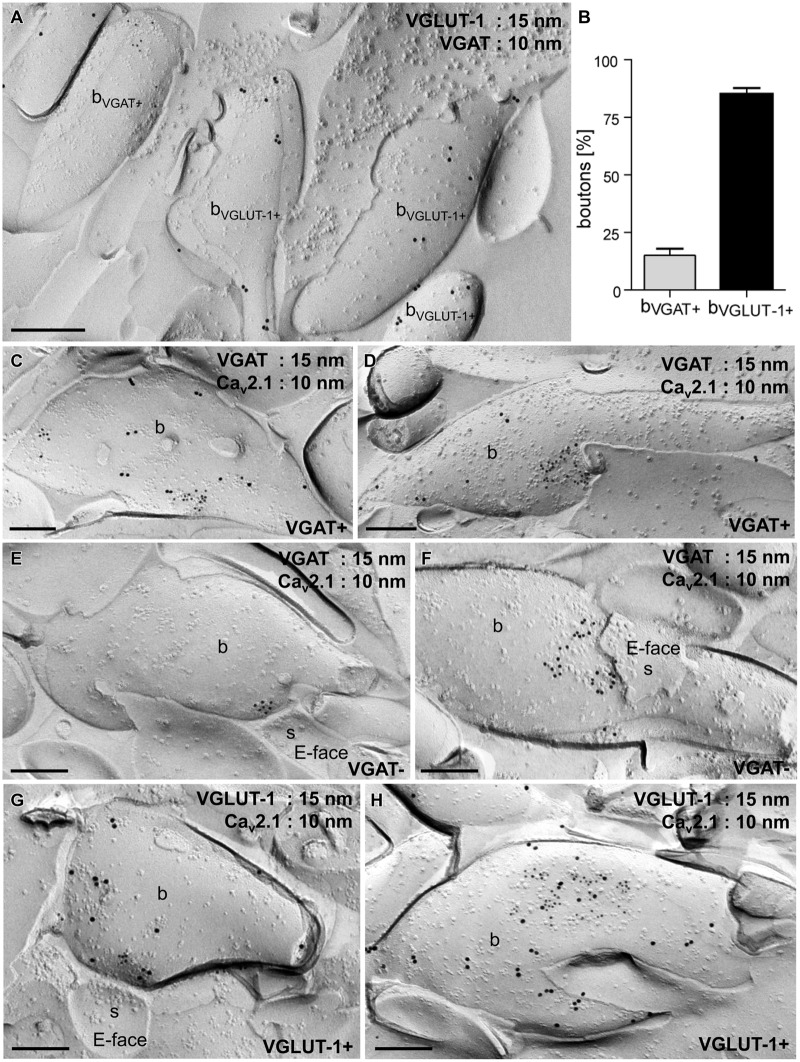
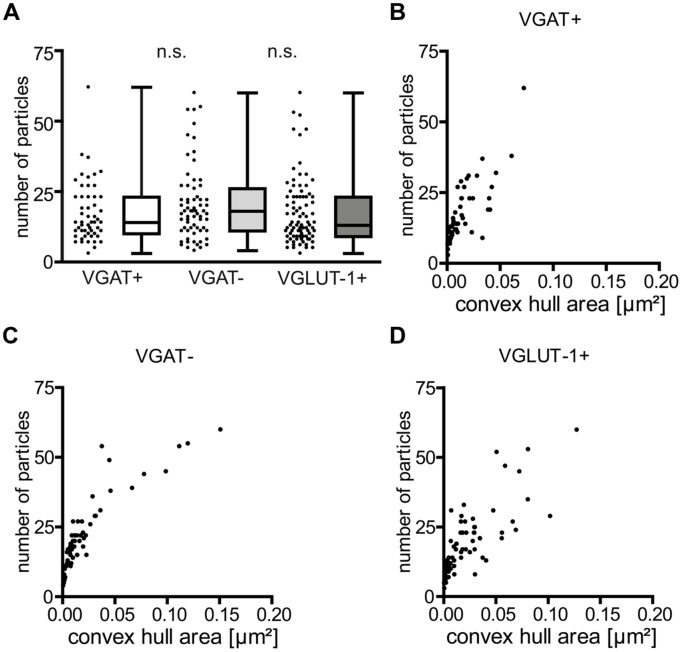
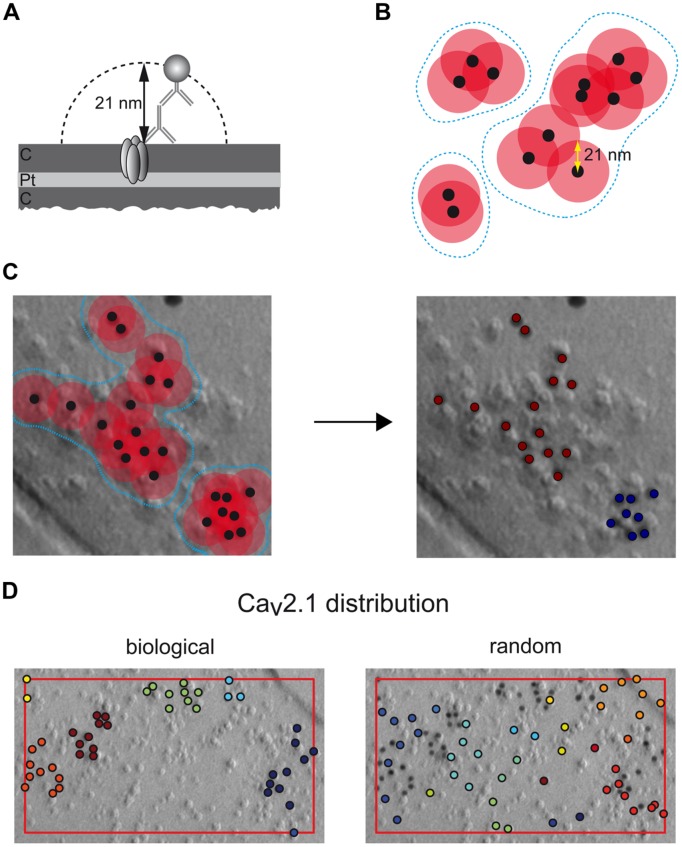
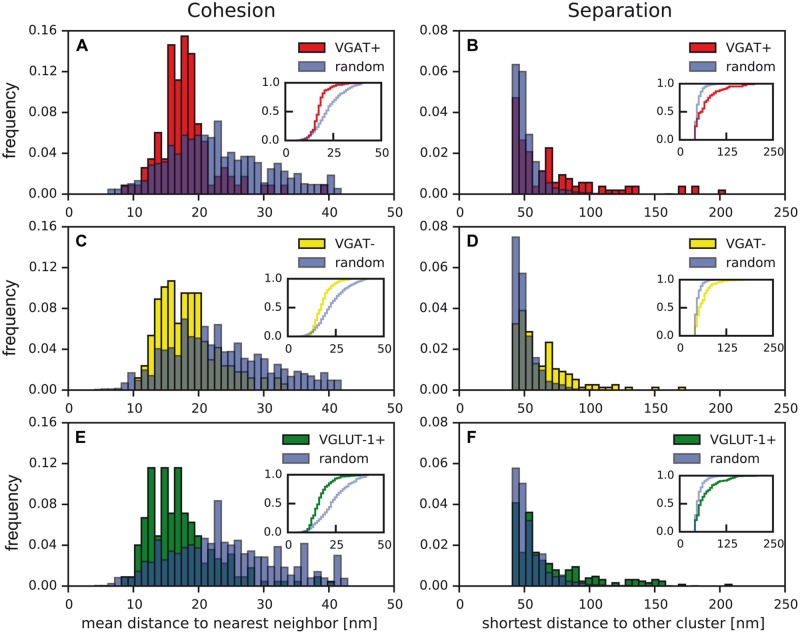
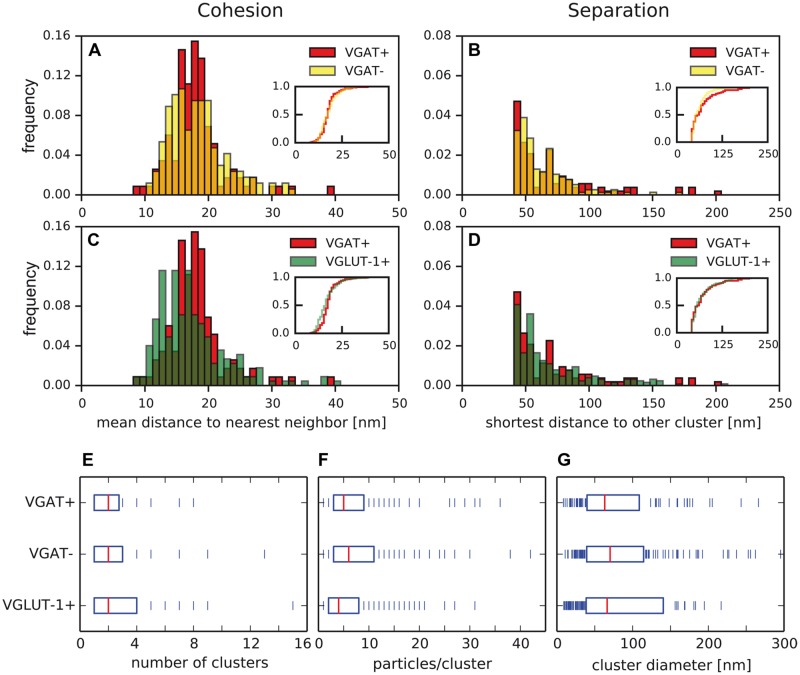
Tuning of the time course and strength of inhibitory and excitatory neurotransmitter release is fundamental for the precise operation of cortical network activity and is controlled by Ca(2+) influx into presynaptic terminals through the high voltage-activated P/Q-type Ca(2+) (Cav2.1) channels. Proper channel-mediated Ca(2+)-signaling critically depends on the topographical arrangement of the channels in the presynaptic membrane. Here, we used high-resolution SDS-digested freeze-fracture replica immunoelectron microscopy together with automatized computational analysis of Cav2.1 immunogold labeling to determine the precise subcellular organization of Cav2.1 channels in both inhibitory and excitatory terminals. Immunoparticles labeling the pore-forming α1 subunit of Cav2.1 channels were enriched over the active zone of the boutons with the number of channels (3-62) correlated with the area of the synaptic membrane. Detailed analysis showed that Cav2.1 channels are non-uniformly distributed over the presynaptic membrane specialization where they are arranged in clusters of an average five channels per cluster covering a mean area with a diameter of about 70 nm. Importantly, clustered arrangement and cluster properties did not show any significant difference between GABAergic and glutamatergic terminals. Our data demonstrate a common nano-architecture of Cav2.1 channels in inhibitory and excitatory boutons in stratum radiatum of the hippocampal CA1 area suggesting that the cluster arrangement is crucial for the precise release of transmitters from the axonal boutons.
Keywords: Ca2+ channels; cluster analysis; hippocampus; quantitative immunoelectron microscopy; rat.
Figures
References
-
- Barber C. B., Dobkin D. P., Huhdanpaa H. (1996). The quickhull algorithm for convex hulls. ACM Trans. Math. Softw. 22 469–483. 10.1145/235815.235821 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous