

The relationship between the neural computations for speech and music perception is context-dependent: an activation likelihood estimate study
- PMID: 26321976
- PMCID: PMC4531212
- DOI: 10.3389/fpsyg.2015.01138
The relationship between the neural computations for speech and music perception is context-dependent: an activation likelihood estimate study
Abstract
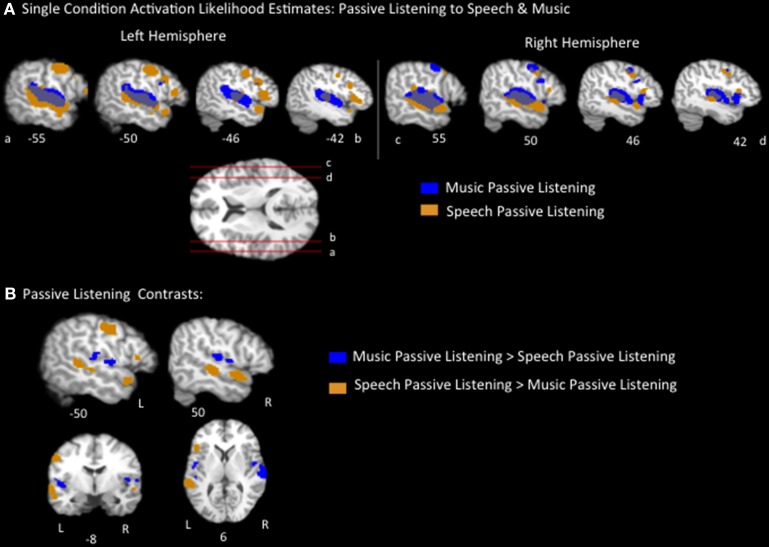
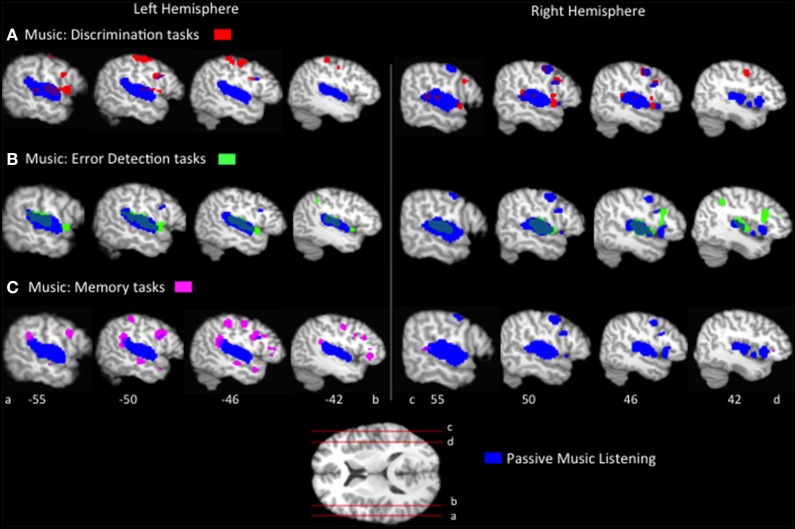
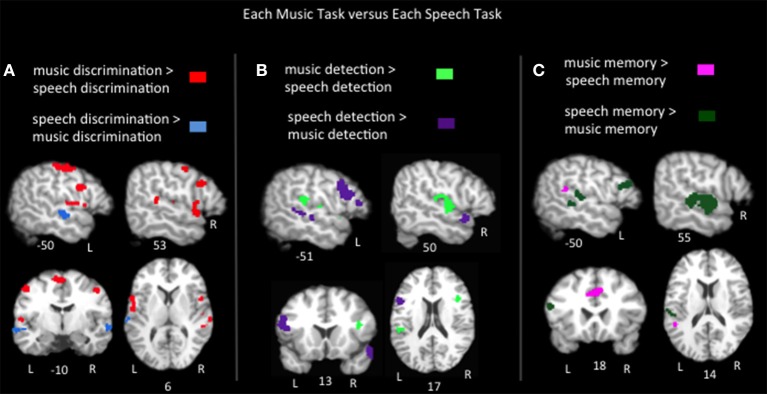
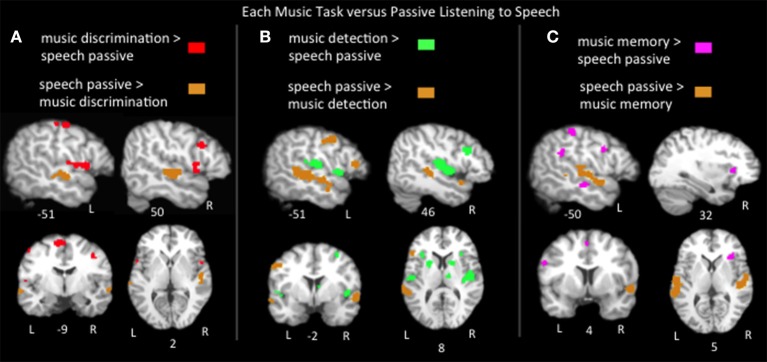
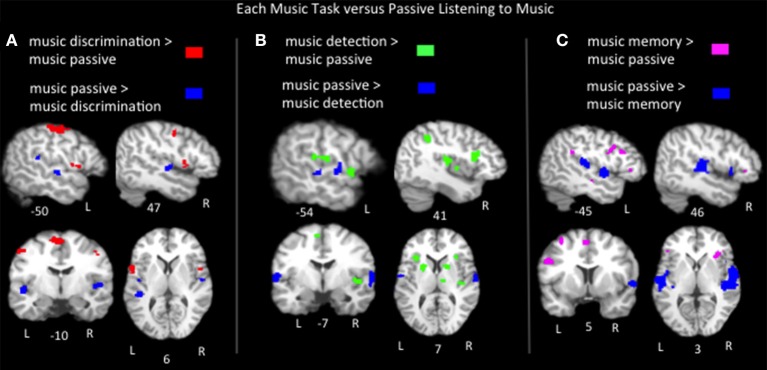
The relationship between the neurobiology of speech and music has been investigated for more than a century. There remains no widespread agreement regarding how (or to what extent) music perception utilizes the neural circuitry that is engaged in speech processing, particularly at the cortical level. Prominent models such as Patel's Shared Syntactic Integration Resource Hypothesis (SSIRH) and Koelsch's neurocognitive model of music perception suggest a high degree of overlap, particularly in the frontal lobe, but also perhaps more distinct representations in the temporal lobe with hemispheric asymmetries. The present meta-analysis study used activation likelihood estimate analyses to identify the brain regions consistently activated for music as compared to speech across the functional neuroimaging (fMRI and PET) literature. Eighty music and 91 speech neuroimaging studies of healthy adult control subjects were analyzed. Peak activations reported in the music and speech studies were divided into four paradigm categories: passive listening, discrimination tasks, error/anomaly detection tasks and memory-related tasks. We then compared activation likelihood estimates within each category for music vs. speech, and each music condition with passive listening. We found that listening to music and to speech preferentially activate distinct temporo-parietal bilateral cortical networks. We also found music and speech to have shared resources in the left pars opercularis but speech-specific resources in the left pars triangularis. The extent to which music recruited speech-activated frontal resources was modulated by task. While there are certainly limitations to meta-analysis techniques particularly regarding sensitivity, this work suggests that the extent of shared resources between speech and music may be task-dependent and highlights the need to consider how task effects may be affecting conclusions regarding the neurobiology of speech and music.
Keywords: Broca's area; fMRI; meta-analysis; music perception; speech perception.
Figures
References
-
- Amunts K., Schleicher A., Bürgel U., Mohlberg H., Uylings H. B. M., Zilles K. (1999). Broca's region revisited: cytoarchitecture and intersubject variability. J. Comp. Neurol. 412, 319–341. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous