

The march toward malaria vaccines
- PMID: 26324116
- PMCID: PMC5634526
- DOI: 10.1016/j.vaccine.2015.07.091
The march toward malaria vaccines
Abstract
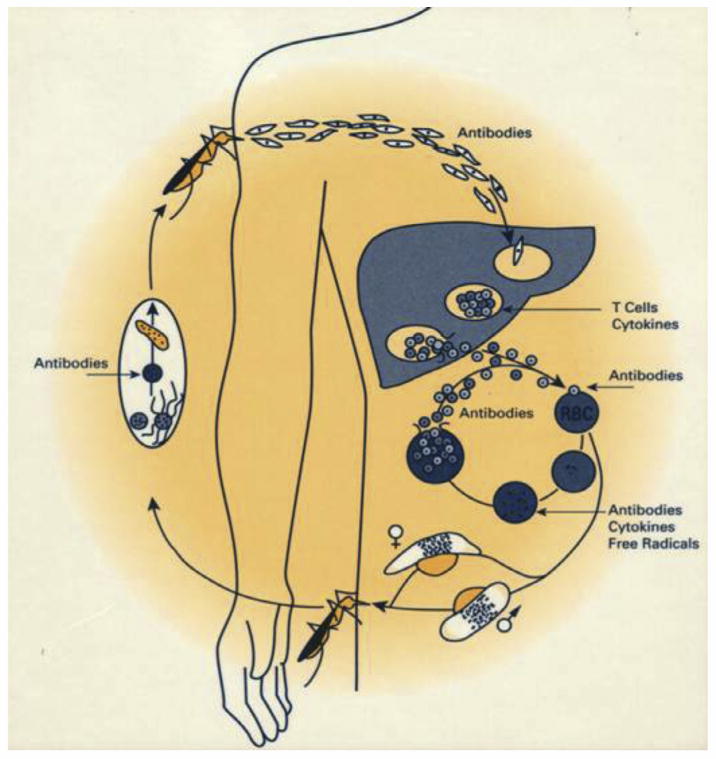
In 2013 there were an estimated 584,000 deaths and 198 million clinical illnesses due to malaria, the majority in sub-Saharan Africa. Vaccines would be the ideal addition to the existing armamentarium of anti-malaria tools. However, malaria is caused by parasites, and parasites are much more complex in terms of their biology than the viruses and bacteria for which we have vaccines, passing through multiple stages of development in the human host, each stage expressing hundreds of unique antigens. This complexity makes it more difficult to develop a vaccine for parasites than for viruses and bacteria, since an immune response targeting one stage may not offer protection against a later stage, because different antigens are the targets of protective immunity at different stages. Furthermore, depending on the life cycle stage and whether the parasite is extra- or intra-cellular, antibody and/or cellular immune responses provide protection. It is thus not surprising that there is no vaccine on the market for prevention of malaria, or any human parasitic infection. In fact, no vaccine for any disease with this breadth of targets and immune responses exists. In this limited review, we focus on four approaches to malaria vaccines, (1) a recombinant protein with adjuvant vaccine aimed at Plasmodium falciparum (Pf) pre-erythrocytic stages of the parasite cycle (RTS,S/AS01), (2) whole sporozoite vaccines aimed at Pf pre-erythrocytic stages (PfSPZ Vaccine and PfSPZ-CVac), (3) prime boost vaccines that include recombinant DNA, viruses and bacteria, and protein with adjuvant aimed primarily at Pf pre-erythrocytic, but also asexual erythrocytic stages, and (4) recombinant protein with adjuvant vaccines aimed at Pf and Plasmodium vivax sexual erythrocytic and mosquito stages. We recognize that we are not covering all approaches to malaria vaccine development, or most of the critically important work on development of vaccines against P. vivax, the second most important cause of malaria. Progress during the last few years has been significant, and a first generation malaria candidate vaccine, RTS,S/AS01, is under review by the European Medicines Agency (EMA) for its quality, safety and efficacy under article 58, which allows the EMA to give a scientific opinion about products intended exclusively for markets outside of the European Union. However, much work is in progress to optimize malaria vaccines in regard to magnitude and durability of protective efficacy and the financing and practicality of delivery. Thus, we are hopeful that anti-malaria vaccines will soon be important tools in the battle against malaria.
Keywords: Malaria; Malaria elimination; Malaria prevention; Malaria vaccines; Plasmodium falciparum; Plasmodium falciparum vaccines; Vaccine.
Copyright © 2015 American Journal of Preventive Medicine. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Figures
References
-
- World Health Organization. World malaria report 2014. Geneva: World Health Organization; 2014.
-
- Hoffman SL, Miller LH. Perspectives on malaria vaccine development. In: Hoffman SL, editor. Malaria vaccine development: a multi-immune response approach. Washington, DC; ASM Press; 1996. pp. 1–13.
-
- Kumar KA, Sano G, Boscardin S, Nussenzweig RS, Nussenzweig MC, Zavala F, et al. The circumsporozoite protein is an immunodominant protective antigen in irradiated sporozoites. Nature. 2006;444:937–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
