

A Matter of Timing: Contrasting Effects of Hydrogen Sulfide on Oxidative Stress Response in Shewanella oneidensis
- PMID: 26324455
- PMCID: PMC4621084
- DOI: 10.1128/JB.00603-15
A Matter of Timing: Contrasting Effects of Hydrogen Sulfide on Oxidative Stress Response in Shewanella oneidensis
Abstract
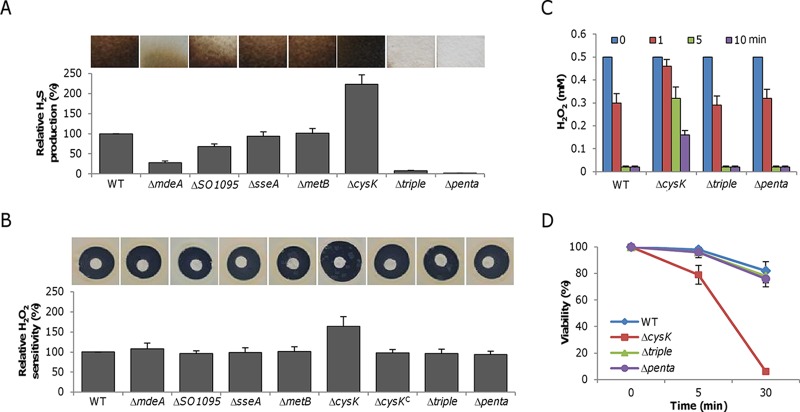
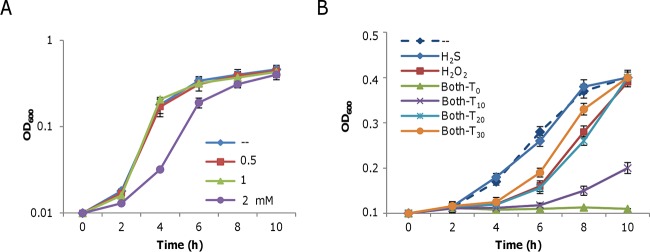
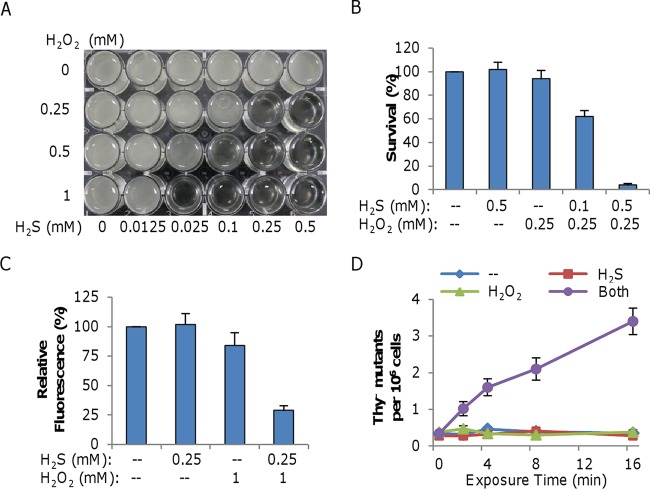
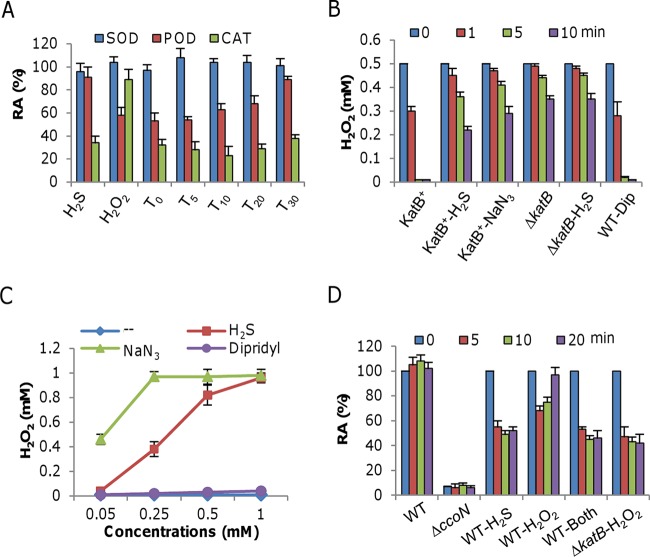
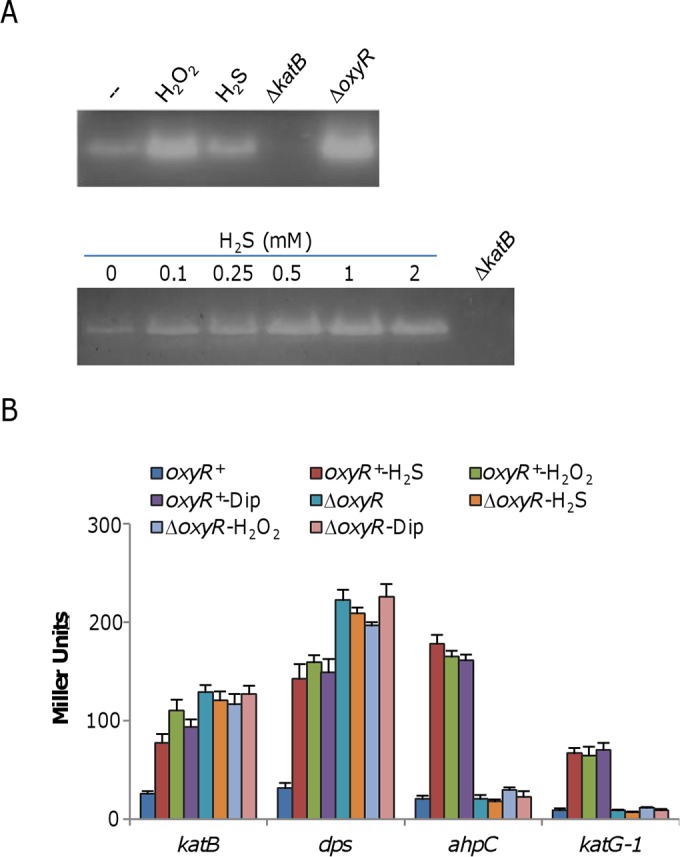
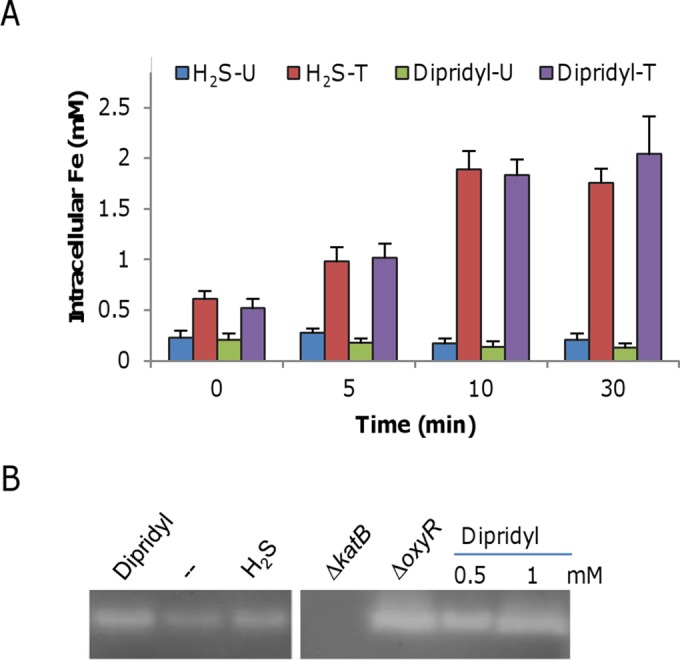
Hydrogen sulfide (H2S), well known for its toxic properties, has recently become a research focus in bacteria, in part because it has been found to prevent oxidative stress caused by treatment with some antibiotics. H2S has the ability to scavenge reactive oxygen species (ROS), thus preventing oxidative stress, but it is also toxic, leading to conflicting reports of its effects in different organisms. Here, with Shewanella oneidensis as a model, we report that the effects of H2S on the response to oxidative stress are time dependent. When added simultaneously with H2O2, H2S promoted H2O2 toxicity by inactivating catalase, KatB, a heme-containing enzyme involved in H2O2 degradation. Such an inhibitory effect may apply to other heme-containing proteins, such as cytochrome cbb3 oxidase. When H2O2 was supplied 20 min or later after the addition of H2S, the oxidative-stress-responding regulator OxyR was activated, resulting in increased resistance to H2O2. The activation of OxyR was likely triggered by the influx of iron, a response to lowered intracellular iron due to the iron-sequestering property of H2S. Given that Shewanella bacteria thrive in redox-stratified environments that have abundant sulfur and iron species, our results imply that H2S is more important for bacterial survival in such environmental niches than previously believed.
Importance: Previous studies have demonstrated that H2S is either detrimental or beneficial to bacterial cells. While it can act as a growth-inhibiting molecule by damaging DNA and denaturing proteins, it helps cells to combat oxidative stress. Here we report that H2S indeed has these contrasting biological functions and that its effects are time dependent. Immediately after H2S treatment, there is growth inhibition due to damage of heme-containing proteins, at least to catalase and cytochrome c oxidase. In contrast, when added a certain time later, H2S confers an enhanced ability to combat oxidative stress by activating the H2O2-responding regulator OxyR. Our data reconcile conflicting observations about the functions of H2S.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
