

Factor XIIIa-dependent retention of red blood cells in clots is mediated by fibrin α-chain crosslinking
- PMID: 26324704
- PMCID: PMC4608241
- DOI: 10.1182/blood-2015-06-652263
Factor XIIIa-dependent retention of red blood cells in clots is mediated by fibrin α-chain crosslinking
Abstract
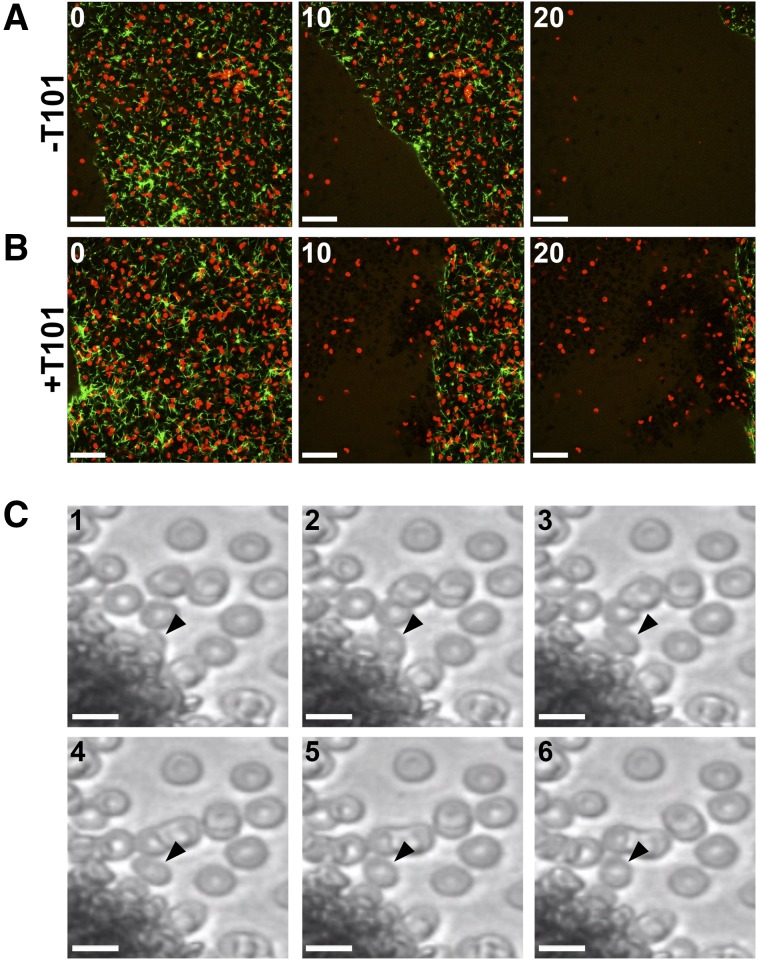
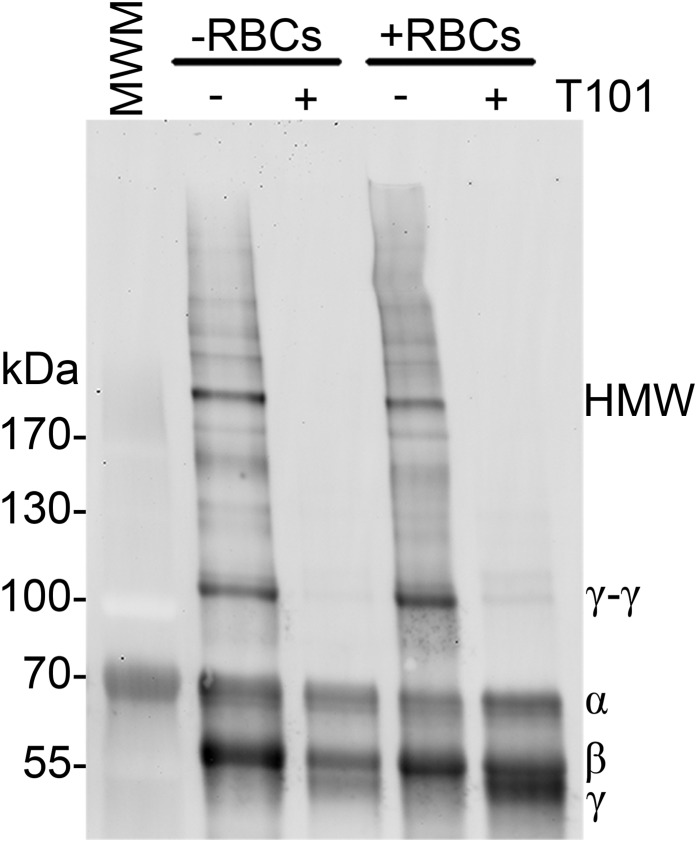
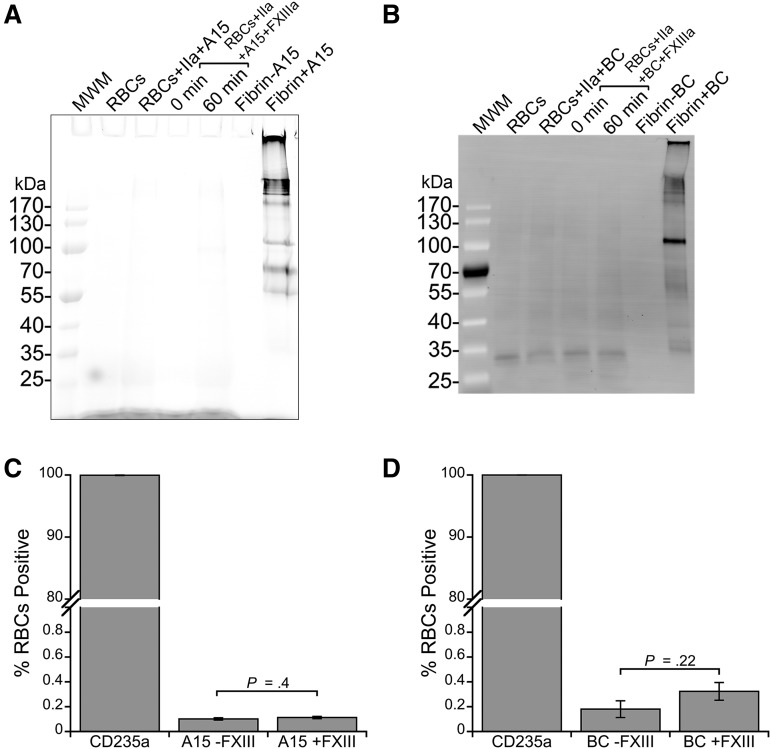
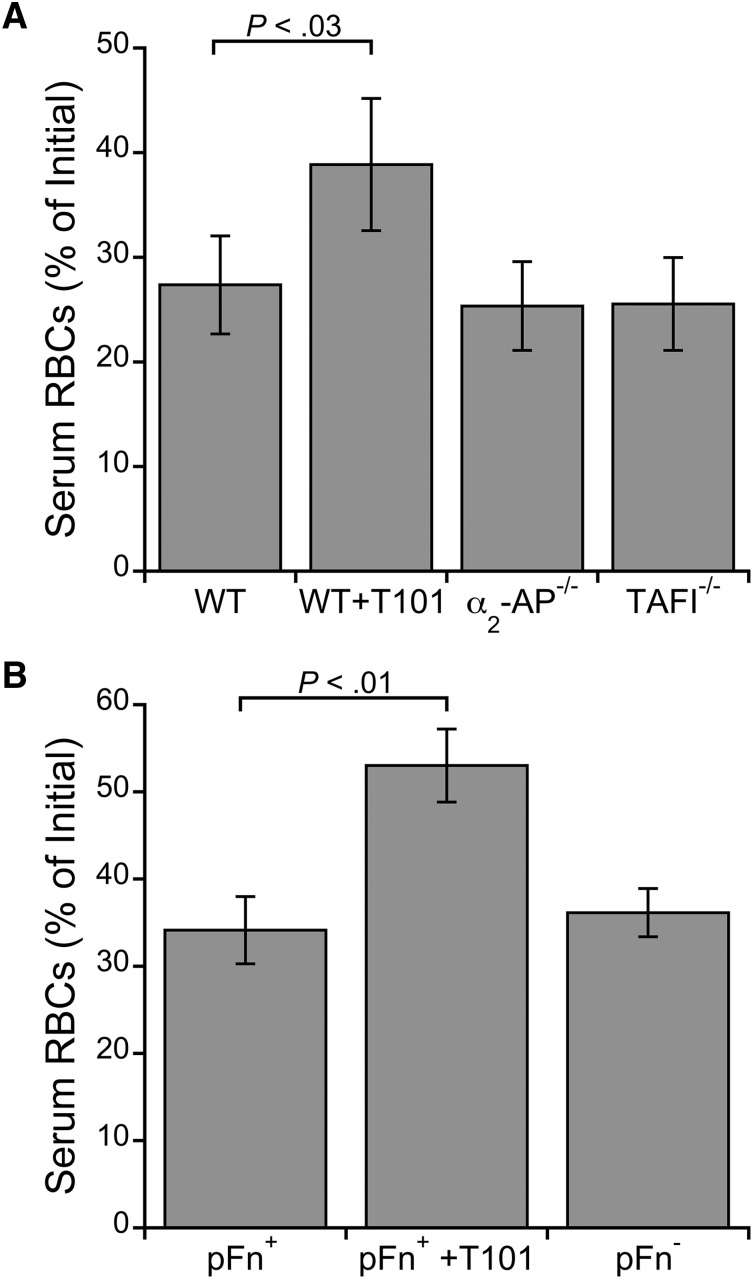
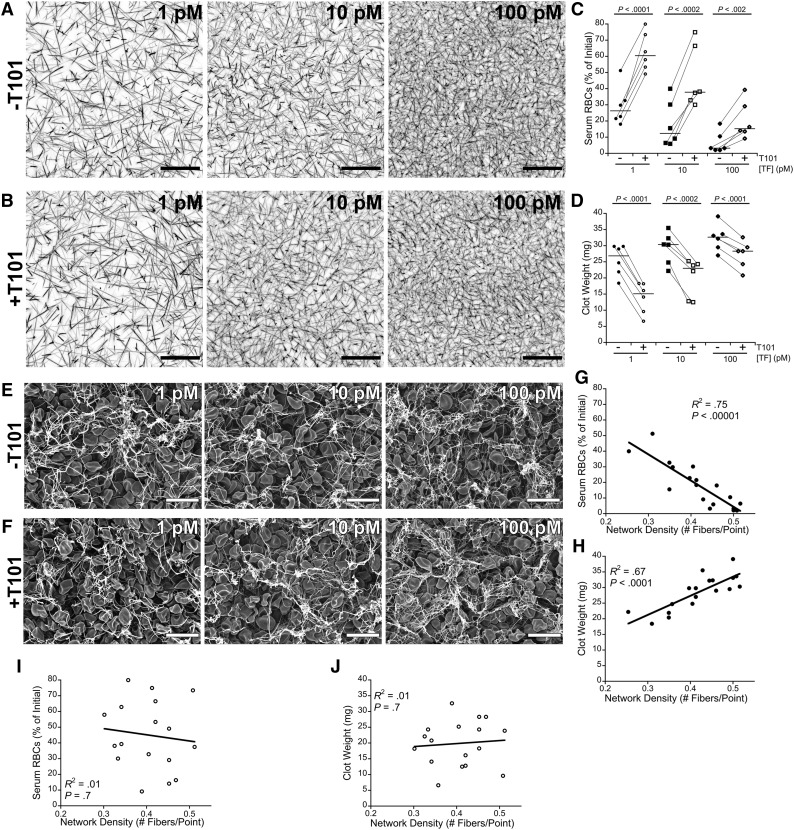
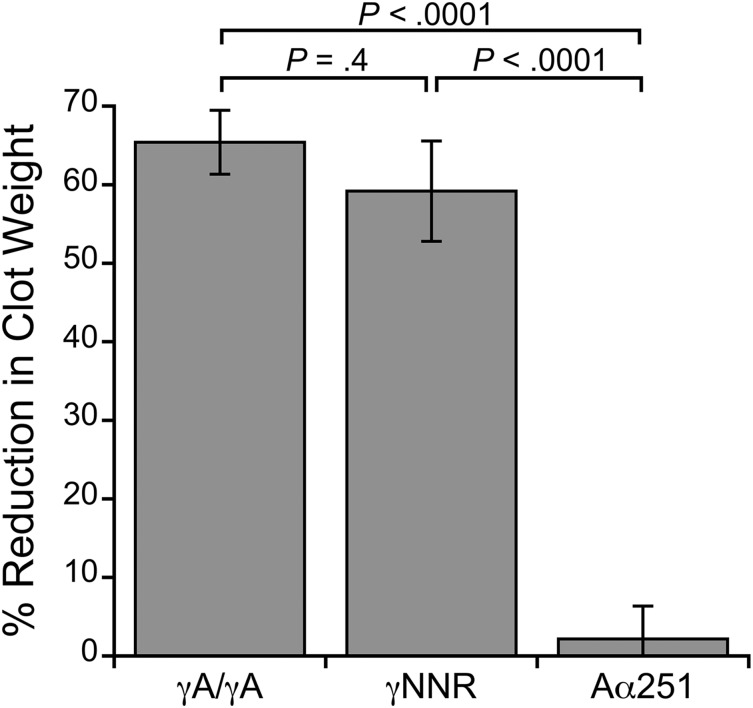
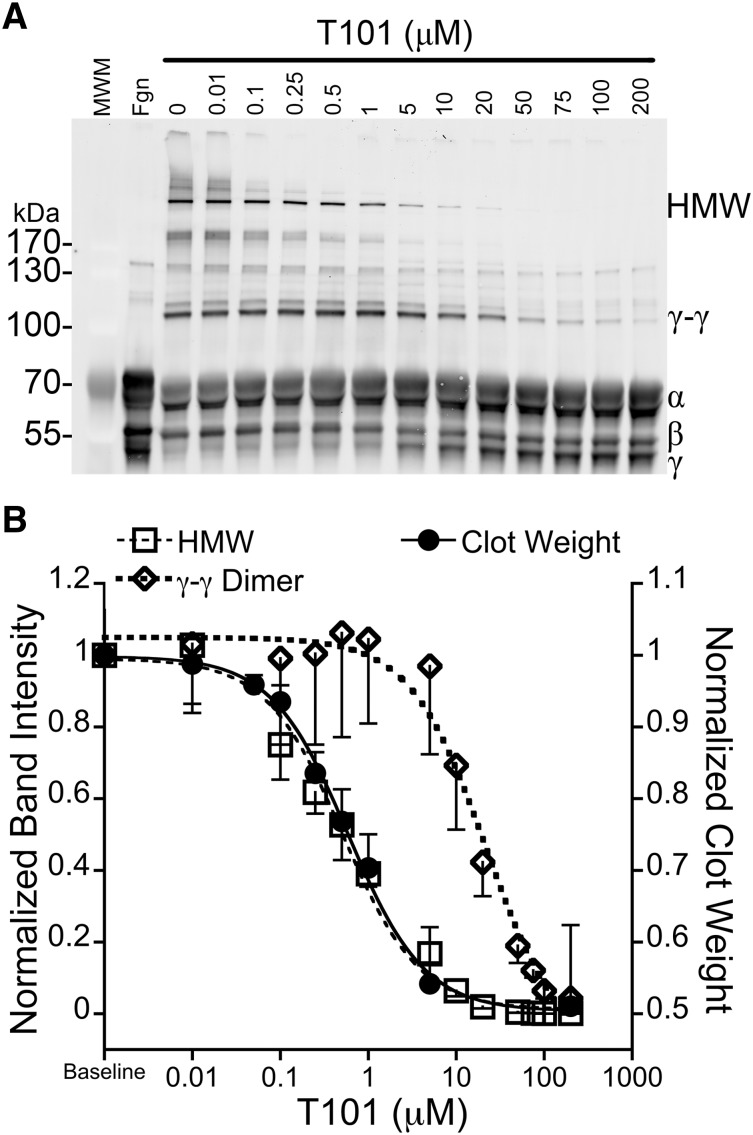
Factor XIII(a) [FXIII(a)] stabilizes clots and increases resistance to fibrinolysis and mechanical disruption. FXIIIa also mediates red blood cell (RBC) retention in contracting clots and determines venous thrombus size, suggesting FXIII(a) is a potential target for reducing thrombosis. However, the mechanism by which FXIIIa retains RBCs in clots is unknown. We determined the effect of FXIII(a) on human and murine clot weight and composition. Real-time microscopy revealed extensive RBC loss from clots formed in the absence of FXIIIa activity, and RBCs exhibited transient deformation as they exited the clots. Fibrin band-shift assays and flow cytometry did not reveal crosslinking of fibrin or FXIIIa substrates to RBCs, suggesting FXIIIa does not crosslink RBCs directly to the clot. RBCs were retained in clots from mice deficient in α2-antiplasmin, thrombin-activatable fibrinolysis inhibitor, or fibronectin, indicating RBC retention does not depend on these FXIIIa substrates. RBC retention in clots was positively correlated with fibrin network density; however, FXIIIa inhibition reduced RBC retention at all network densities. FXIIIa inhibition reduced RBC retention in clots formed with fibrinogen that lacks γ-chain crosslinking sites, but not in clots that lack α-chain crosslinking sites. Moreover, FXIIIa inhibitor concentrations that primarily block α-, but not γ-, chain crosslinking decreased RBC retention in clots. These data indicate FXIIIa-dependent retention of RBCs in clots is mediated by fibrin α-chain crosslinking. These findings expose a newly recognized, essential role for fibrin crosslinking during whole blood clot formation and consolidation and establish FXIIIa activity as a key determinant of thrombus composition and size.
© 2015 by The American Society of Hematology.
Figures
References
-
- Lak M, Keihani M, Elahi F, Peyvandi F, Mannucci PM. Bleeding and thrombosis in 55 patients with inherited afibrinogenaemia. Br J Haematol. 1999;107(1):204–206. - PubMed
-
- Wilhelmsen L, Svärdsudd K, Korsan-Bengtsen K, Larsson B, Welin L, Tibblin G. Fibrinogen as a risk factor for stroke and myocardial infarction. N Engl J Med. 1984;311(8):501–505. - PubMed
-
- Kamphuisen PW, Eikenboom JC, Vos HL, et al. Increased levels of factor VIII and fibrinogen in patients with venous thrombosis are not caused by acute phase reactions. Thromb Haemost. 1999;81(5):680–683. - PubMed
-
- Koster T, Rosendaal FR, Reitsma PH, van der Velden PA, Briët E, Vandenbroucke JP. Factor VII and fibrinogen levels as risk factors for venous thrombosis. A case-control study of plasma levels and DNA polymorphisms--the Leiden Thrombophilia Study (LETS). Thromb Haemost. 1994;71(6):719–722. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
