

Failure of lysosome clustering and positioning in the juxtanuclear region in cells deficient in rapsyn
- PMID: 26330529
- PMCID: PMC4631781
- DOI: 10.1242/jcs.172536
Failure of lysosome clustering and positioning in the juxtanuclear region in cells deficient in rapsyn
Abstract
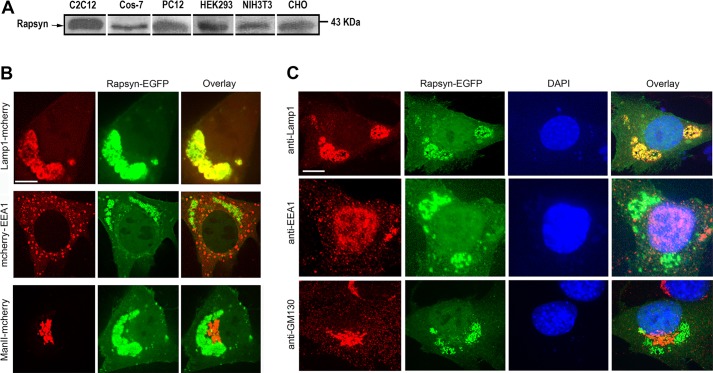
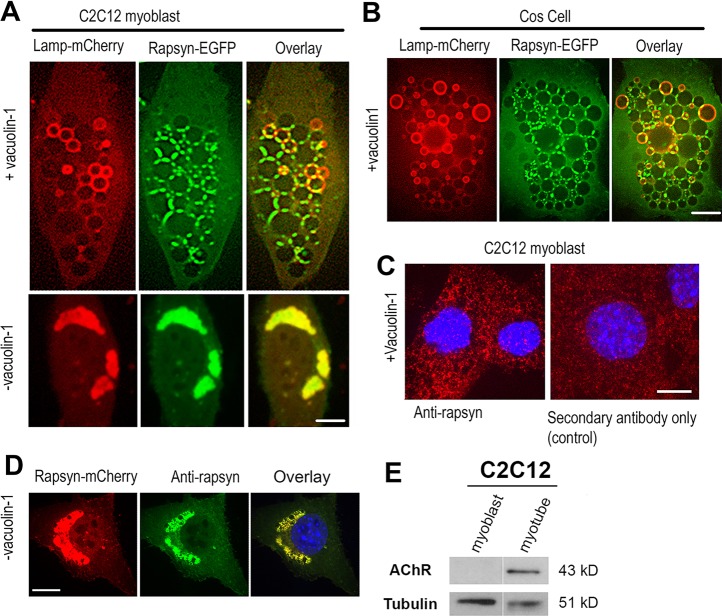
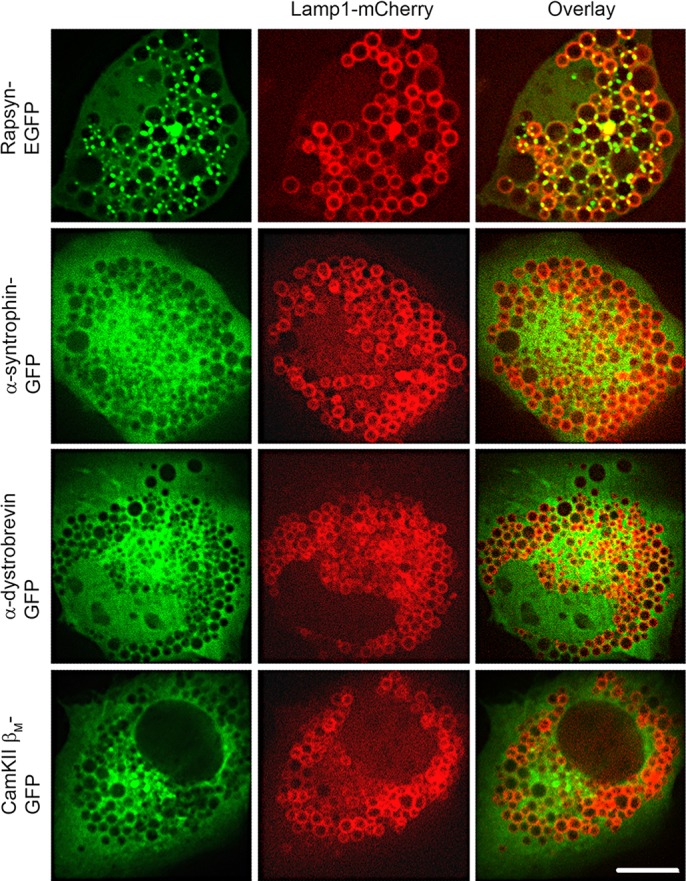
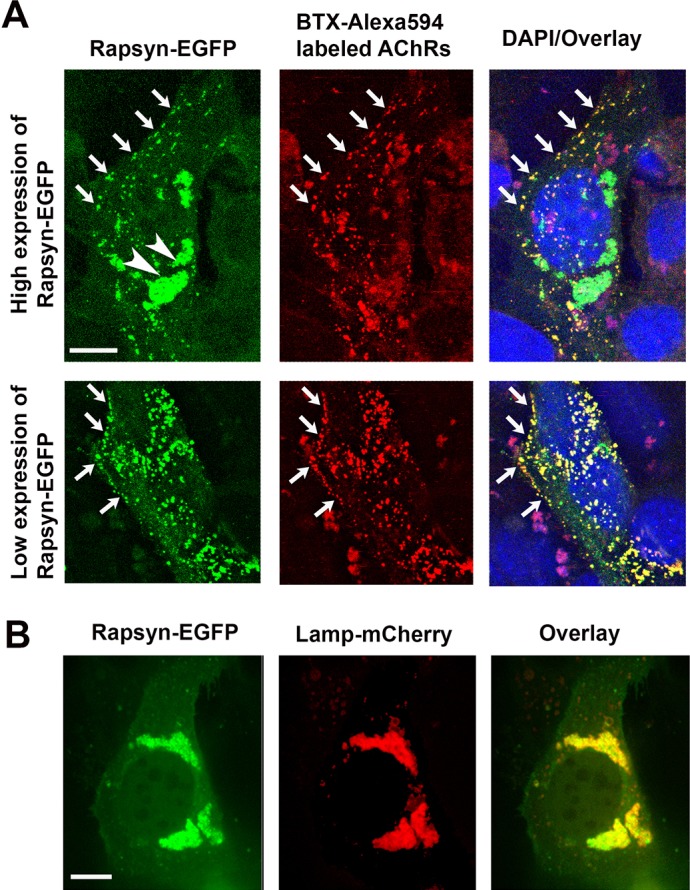
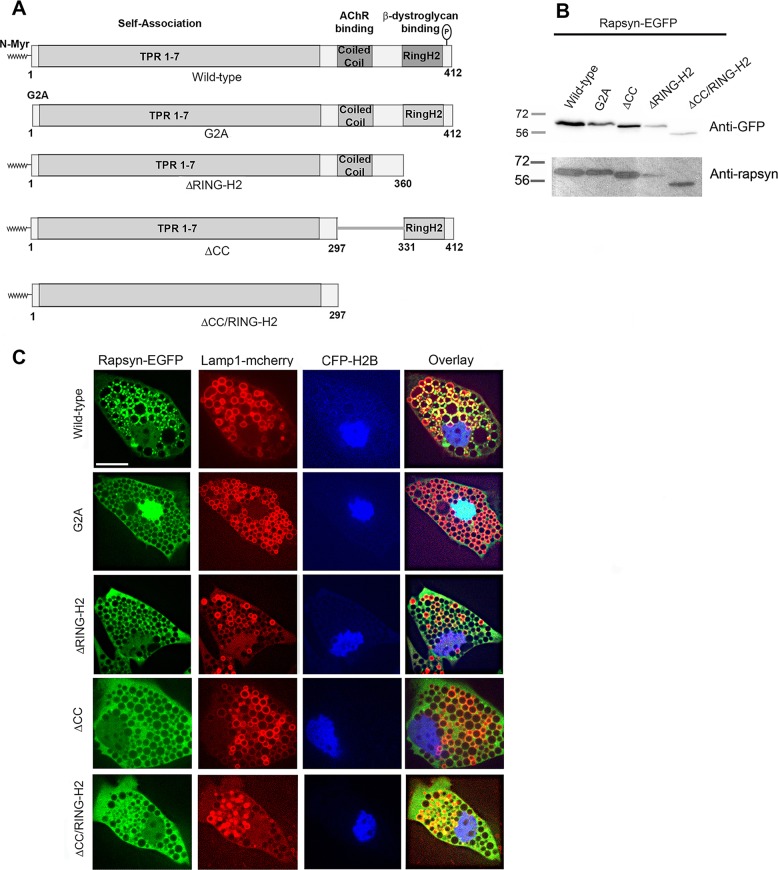
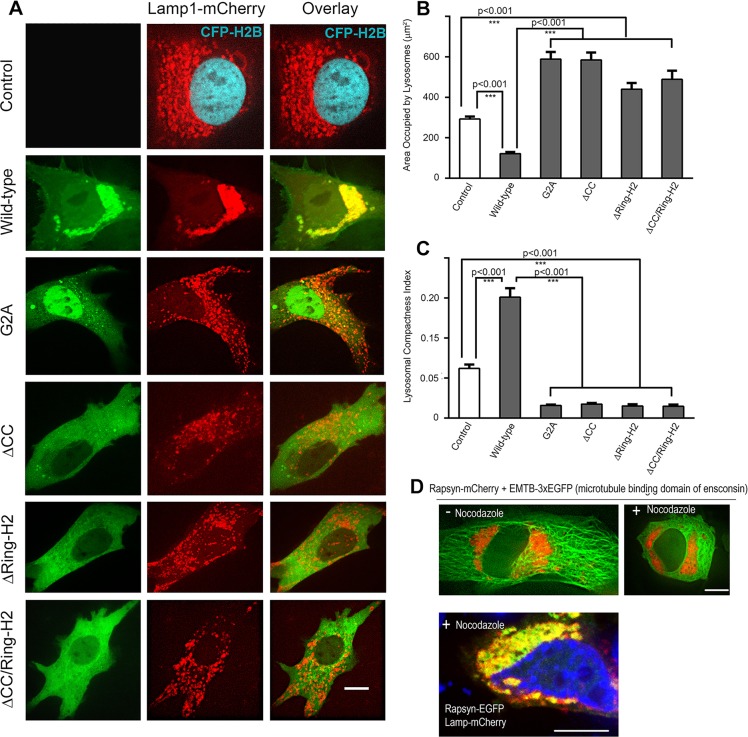
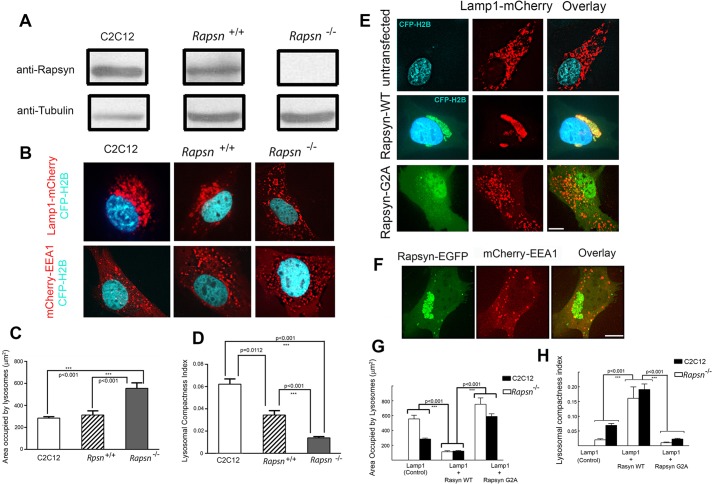
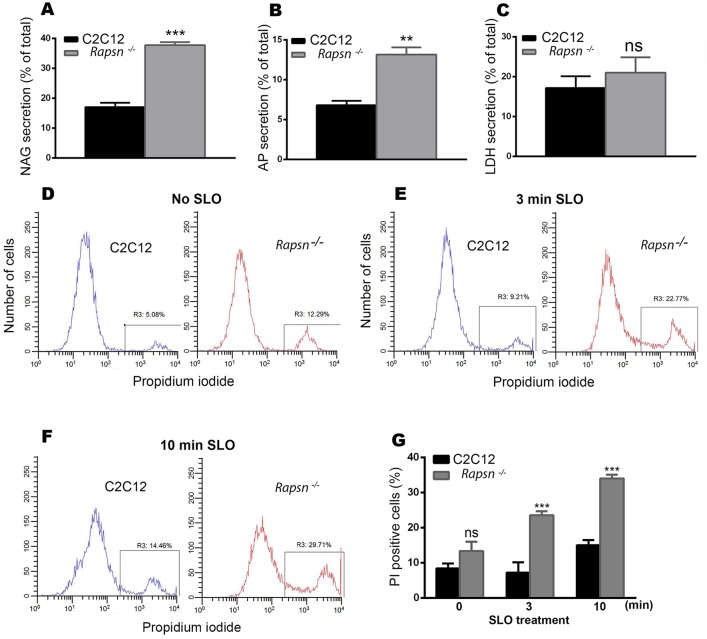
Rapsyn, a scaffold protein, is required for the clustering of acetylcholine receptors (AChRs) at contacts between motor neurons and differentiating muscle cells. Rapsyn is also expressed in cells that do not express AChRs. However, its function in these cells remains unknown. Here, we show that rapsyn plays an AChR-independent role in organizing the distribution and mobility of lysosomes. In cells devoid of AChRs, rapsyn selectively induces the clustering of lysosomes at high density in the juxtanuclear region without affecting the distribution of other intracellular organelles. However, when the same cells overexpress AChRs, rapsyn is recruited away from lysosomes to colocalize with AChR clusters on the cell surface. In rapsyn-deficient (Rapsn(-/-)) myoblasts or cells overexpressing rapsyn mutants, lysosomes are scattered within the cell and highly dynamic. The increased mobility of lysosomes in Rapsn(-/-) cells is associated with a significant increase in lysosomal exocytosis, as evidenced by increased release of lysosomal enzymes and plasma membrane damage when cells were challenged with the bacterial pore-forming toxin streptolysin-O. These findings uncover a new link between rapsyn, lysosome positioning, exocytosis and plasma membrane integrity.
Keywords: Clustering; Exocytosis; Lysosomes; Mobility; Rapsyn.
© 2015. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
