

Protein-Protein Interactions Modulate the Docking-Dependent E3-Ubiquitin Ligase Activity of Carboxy-Terminus of Hsc70-Interacting Protein (CHIP)
- PMID: 26330542
- PMCID: PMC4638040
- DOI: 10.1074/mcp.M115.051169
Protein-Protein Interactions Modulate the Docking-Dependent E3-Ubiquitin Ligase Activity of Carboxy-Terminus of Hsc70-Interacting Protein (CHIP)
Abstract
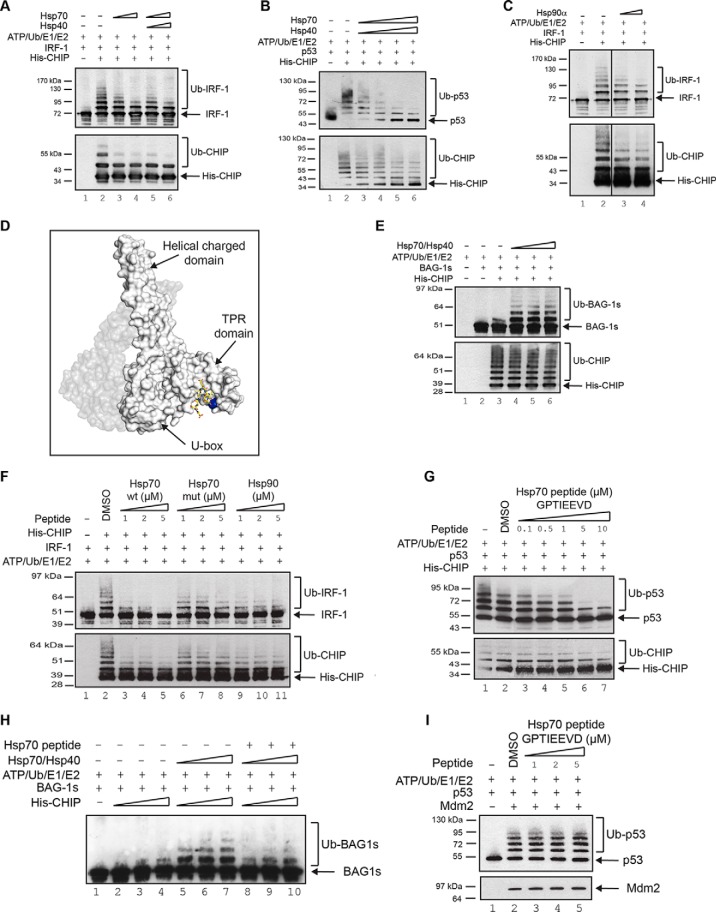
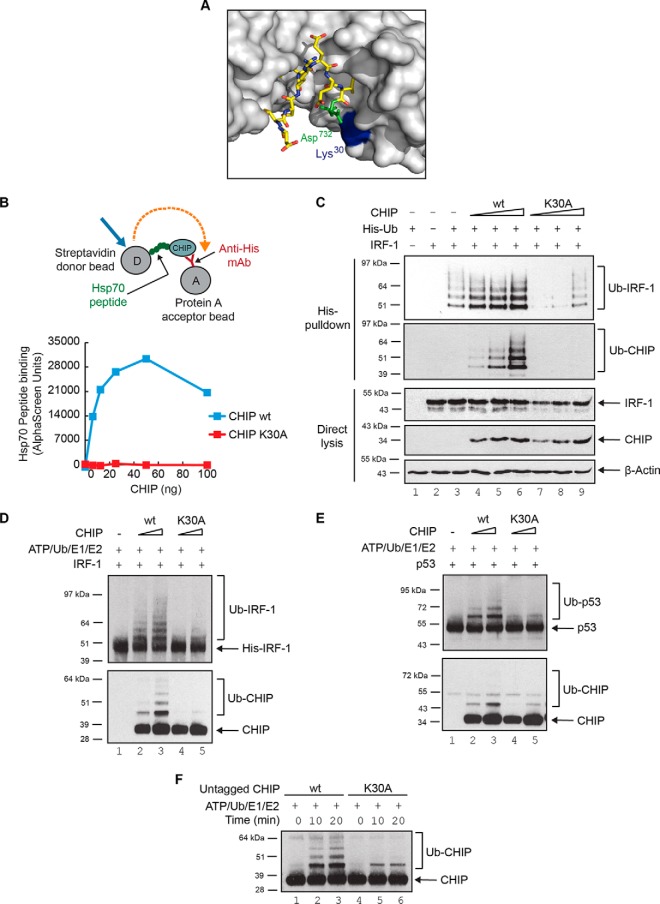
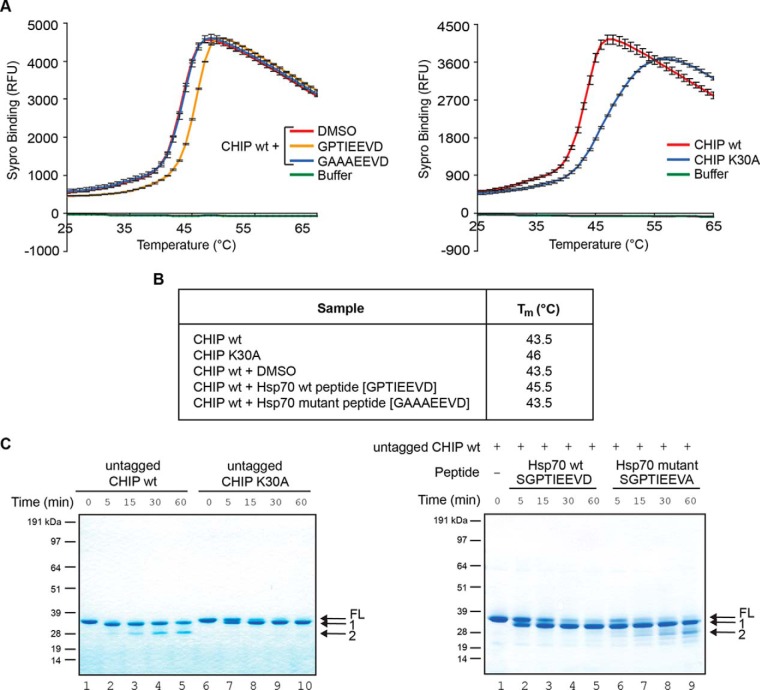
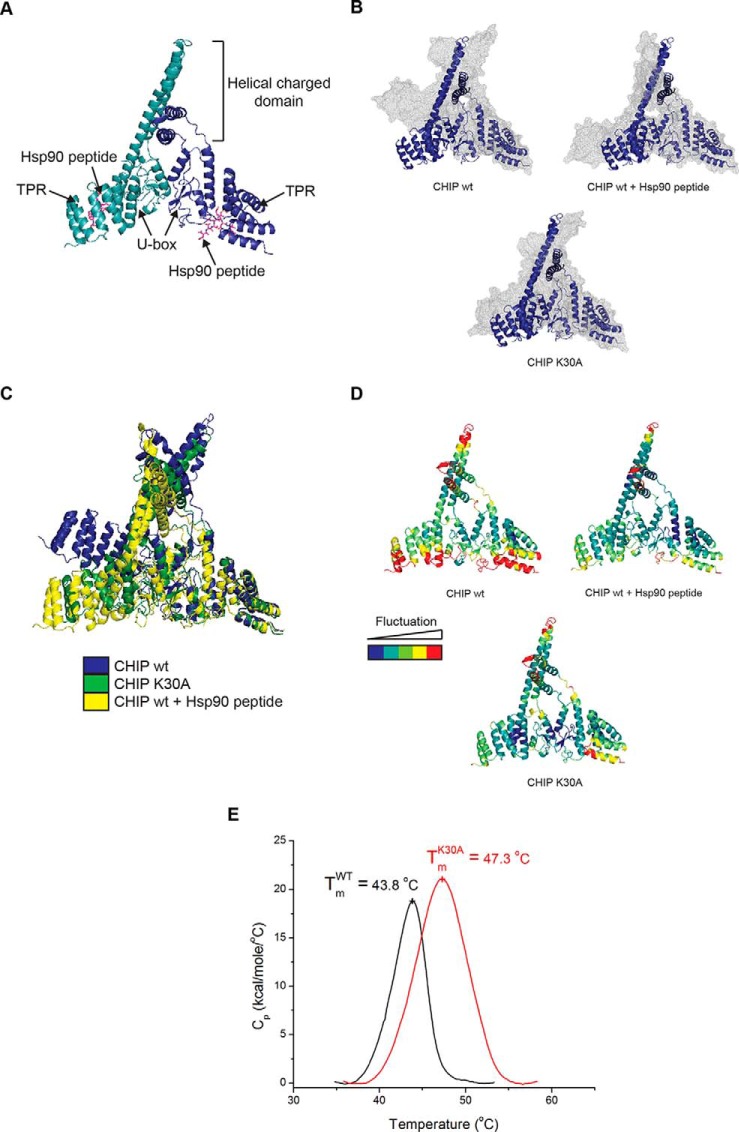
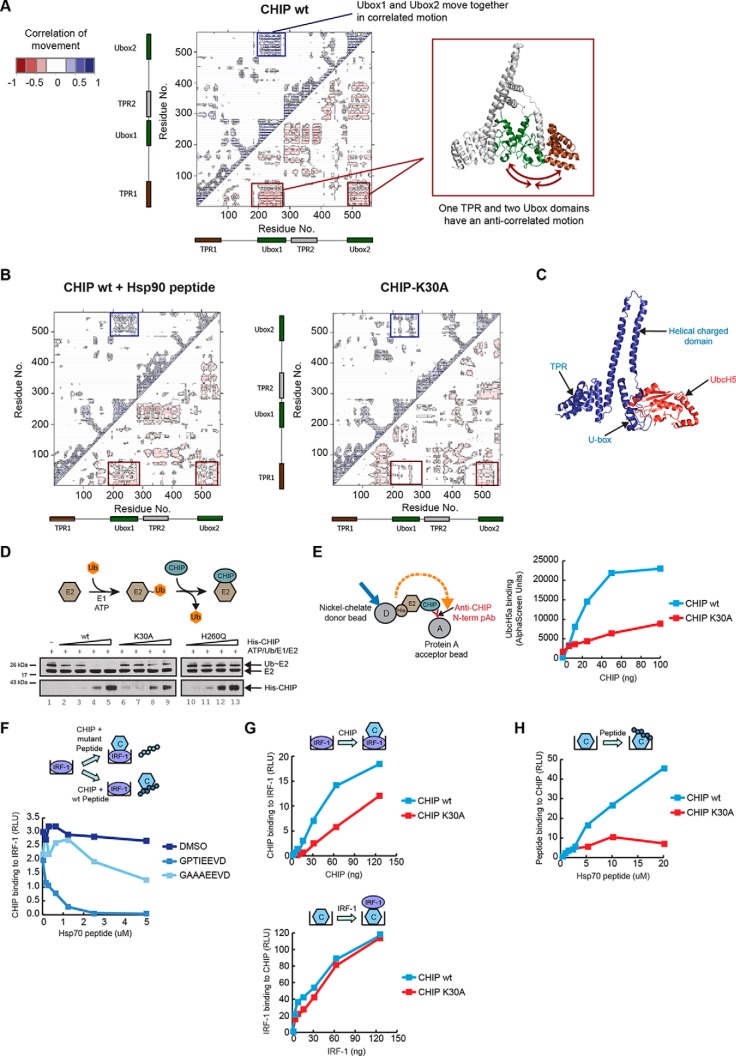
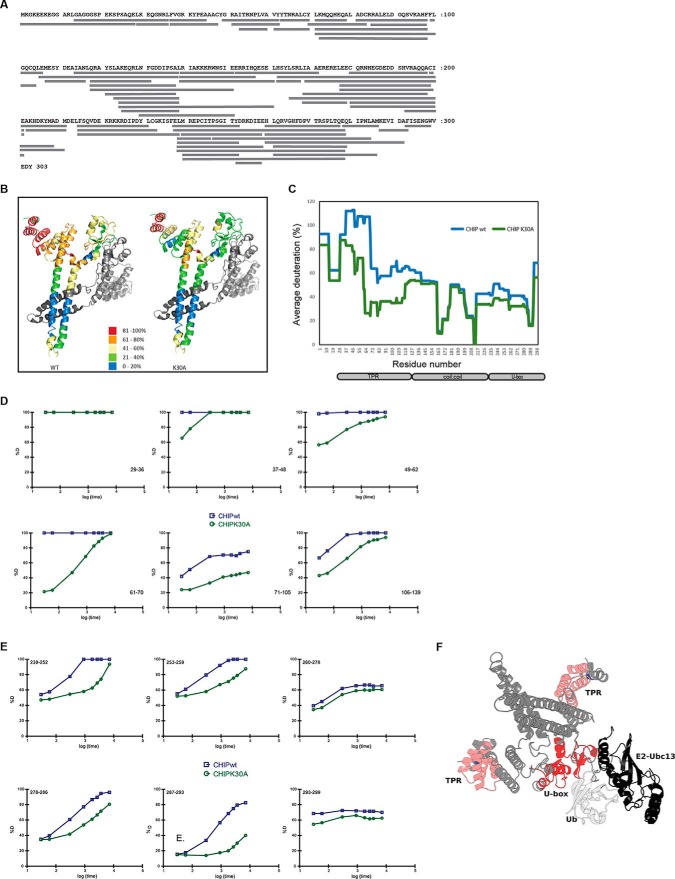
CHIP is a tetratricopeptide repeat (TPR) domain protein that functions as an E3-ubiquitin ligase. As well as linking the molecular chaperones to the ubiquitin proteasome system, CHIP also has a docking-dependent mode where it ubiquitinates native substrates, thereby regulating their steady state levels and/or function. Here we explore the effect of Hsp70 on the docking-dependent E3-ligase activity of CHIP. The TPR-domain is revealed as a binding site for allosteric modulators involved in determining CHIP's dynamic conformation and activity. Biochemical, biophysical and modeling evidence demonstrate that Hsp70-binding to the TPR, or Hsp70-mimetic mutations, regulate CHIP-mediated ubiquitination of p53 and IRF-1 through effects on U-box activity and substrate binding. HDX-MS was used to establish that conformational-inhibition-signals extended from the TPR-domain to the U-box. This underscores inter-domain allosteric regulation of CHIP by the core molecular chaperones. Defining the chaperone-associated TPR-domain of CHIP as a manager of inter-domain communication highlights the potential for scaffolding modules to regulate, as well as assemble, complexes that are fundamental to protein homeostatic control.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- D'Andrea L.D., and Regan L. (2003) TPR proteins: The versatile helix. Trends Biochem. Sci. 28, 655–662 - PubMed
-
- Zeytuni N., and Zarivach R. (2012) Structural and functional discussion of the tetra-trico-peptide repeat, a protein interaction module. Structure 20, 397–405 - PubMed
-
- Andrade M. A., Perez-Iratxeta C., and Ponting C.P. (2001) Protein repeats: structures, functions, and evolution. J. Struct. Biol. 134, 117–131 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
