

CD73-mediated adenosine production promotes stem cell-like properties in mouse Tc17 cells
- PMID: 26331349
- PMCID: PMC4693903
- DOI: 10.1111/imm.12529
CD73-mediated adenosine production promotes stem cell-like properties in mouse Tc17 cells
Abstract
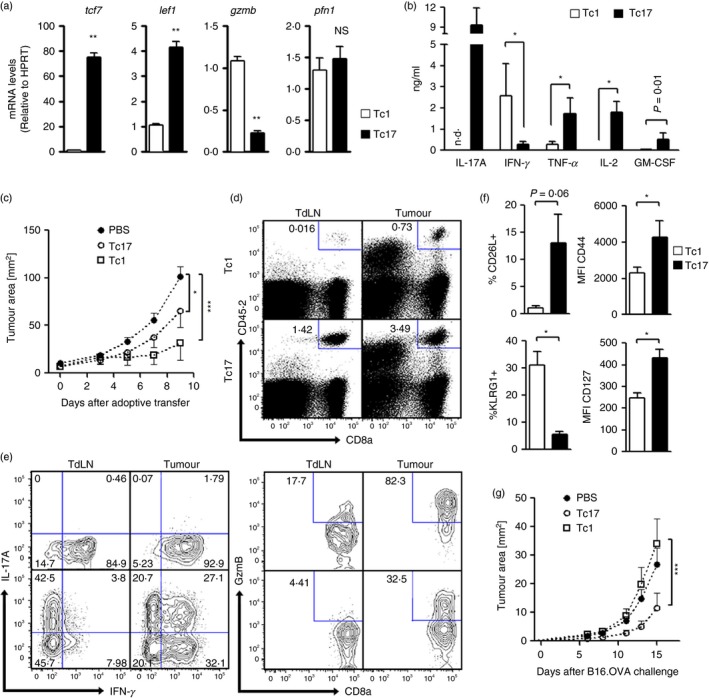
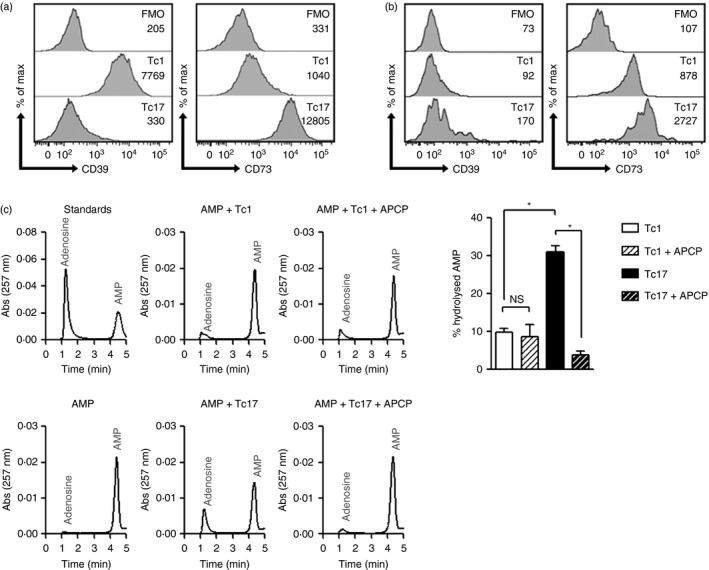
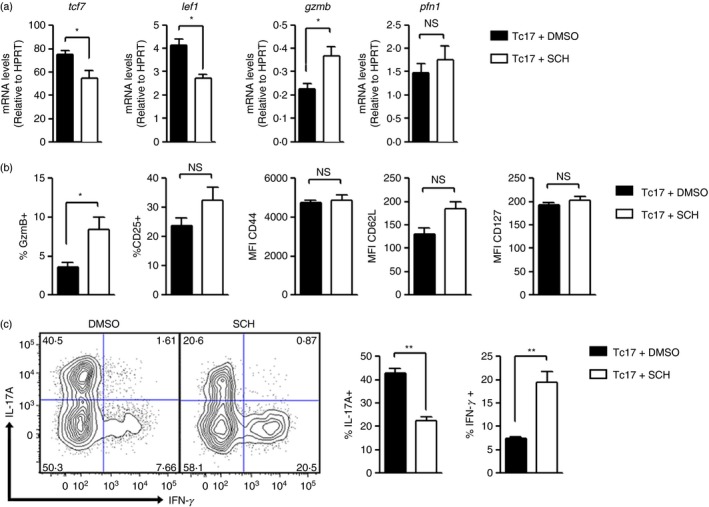
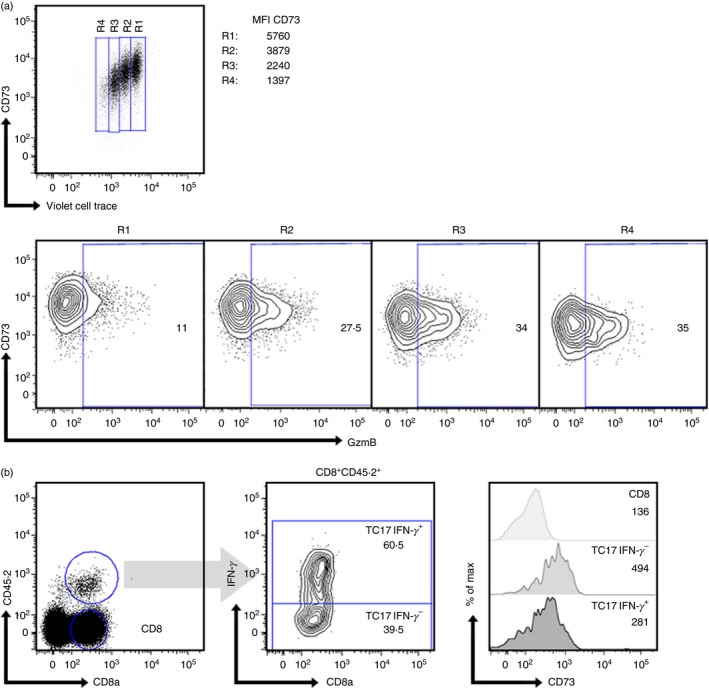
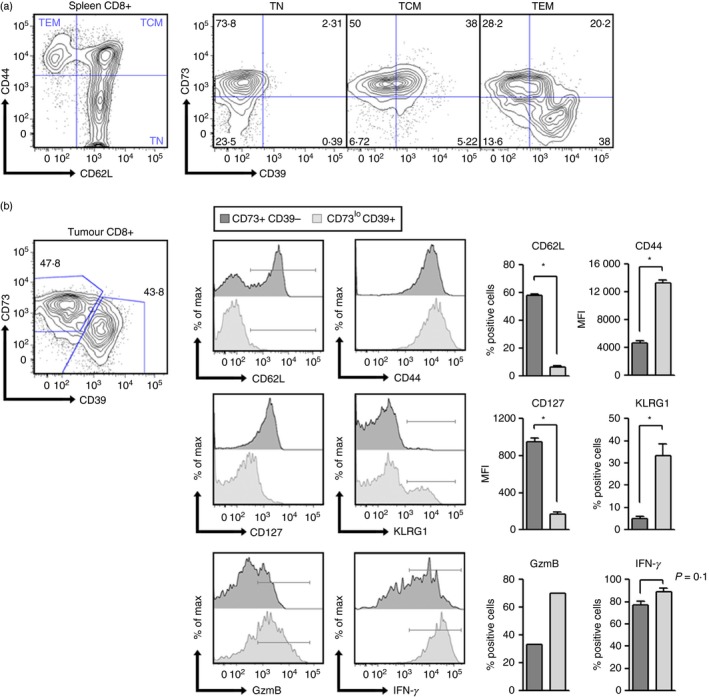
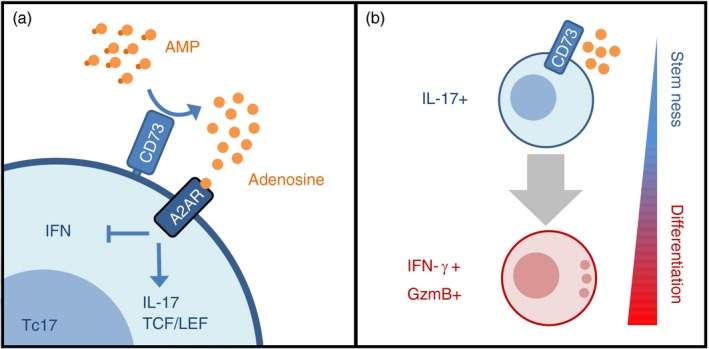
The CD73 ectonucleotidase catalyses the hydrolysis of AMP to adenosine, an immunosuppressive molecule. Recent evidence has demonstrated that this ectonucleotidase is up-regulated in T helper type 17 cells when generated in the presence of transforming growth factor-β (TGF-β), and hence CD73 expression is related to the acquisition of immunosuppressive potential by these cells. TGF-β is also able to induce CD73 expression in CD8(+) T cells but the function of this ectonucleotidase in CD8(+) T cells is still unknown. Here, we show that Tc17 cells present high levels of the CD73 ectonucleotidase and produce adenosine; however, they do not suppress the proliferation of CD4(+) T cells. Interestingly, we report that adenosine signalling through A2A receptor favours interleukin-17 production and the expression of stem cell-associated transcription factors such as tcf-7 and lef-1 but restrains the acquisition of Tc1-related effector molecules such as interferon-γ and Granzyme B by Tc17 cells. Within the tumour microenvironment, CD73 is highly expressed in CD62L(+) CD127(+) CD8(+) T cells (memory T cells) and is down-regulated in GZMB(+) KLRG1(+) CD8(+) T cells (terminally differentiated T cells), demonstrating that CD73 is expressed in memory/naive cells and is down-regulated during differentiation. These data reveal a novel function of CD73 ectonucleotidase in arresting CD8(+) T-cell differentiation and support the idea that CD73-driven adenosine production by Tc17 cells may promote stem cell-like properties in Tc17 cells.
Keywords: CD73; Tc17; adenosine; cell differentiation; stem cell; tumour immunology.
© 2015 John Wiley & Sons Ltd.
Figures
Similar articles
-
Decreased frequency of CD73+CD8+ T cells of HIV-infected patients correlates with immune activation and T cell exhaustion.J Leukoc Biol. 2013 Oct;94(4):551-61. doi: 10.1189/jlb.0113018. Epub 2013 May 24. J Leukoc Biol. 2013. PMID: 23709688
-
Targeting CD73 enhances the antitumor activity of anti-PD-1 and anti-CTLA-4 mAbs.Clin Cancer Res. 2013 Oct 15;19(20):5626-35. doi: 10.1158/1078-0432.CCR-13-0545. Epub 2013 Aug 27. Clin Cancer Res. 2013. PMID: 23983257
-
Generation of anti-inflammatory adenosine by leukocytes is regulated by TGF-β.Eur J Immunol. 2011 Oct;41(10):2955-65. doi: 10.1002/eji.201141512. Epub 2011 Sep 6. Eur J Immunol. 2011. PMID: 21770045
-
Heterogeneity in the differentiation and function of CD8⁺ T cells.Arch Immunol Ther Exp (Warsz). 2014 Dec;62(6):449-58. doi: 10.1007/s00005-014-0293-y. Epub 2014 May 31. Arch Immunol Ther Exp (Warsz). 2014. PMID: 24879097 Review.
-
CD73 and CD39 ectonucleotidases in T cell differentiation: Beyond immunosuppression.FEBS Lett. 2015 Nov 14;589(22):3454-60. doi: 10.1016/j.febslet.2015.07.027. Epub 2015 Jul 29. FEBS Lett. 2015. PMID: 26226423 Review.
Cited by
-
Crosstalk Between ATP-P2X7 and Adenosine A2A Receptors Controlling Neuroinflammation in Rats Subject to Repeated Restraint Stress.Front Cell Neurosci. 2021 Mar 1;15:639322. doi: 10.3389/fncel.2021.639322. eCollection 2021. Front Cell Neurosci. 2021. PMID: 33732112 Free PMC article.
-
CD73 expression identifies a subset of IgM+ antigen-experienced cells with memory attributes that is T cell and CD40 signalling dependent.Immunology. 2017 Dec;152(4):602-612. doi: 10.1111/imm.12800. Epub 2017 Aug 23. Immunology. 2017. PMID: 28746783 Free PMC article.
-
The Multifaceted Actions of CD73 During Development and Suppressive Actions of Regulatory T Cells.Front Immunol. 2022 May 31;13:914799. doi: 10.3389/fimmu.2022.914799. eCollection 2022. Front Immunol. 2022. PMID: 35711418 Free PMC article. Review.
-
Inhibition of the adenosinergic pathway: the indispensable part of oncological therapy in the future.Purinergic Signal. 2019 Mar;15(1):53-67. doi: 10.1007/s11302-018-9641-4. Epub 2019 Feb 26. Purinergic Signal. 2019. PMID: 30809739 Free PMC article. Review.
-
The yin and yang functions of extracellular ATP and adenosine in tumor immunity.Cancer Cell Int. 2020 Apr 7;20:110. doi: 10.1186/s12935-020-01195-x. eCollection 2020. Cancer Cell Int. 2020. PMID: 32280302 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
