

NETosis in Neonates: Evidence of a Reactive Oxygen Species-Independent Pathway in Response to Fungal Challenge
- PMID: 26333942
- PMCID: PMC4721906
- DOI: 10.1093/infdis/jiv435
NETosis in Neonates: Evidence of a Reactive Oxygen Species-Independent Pathway in Response to Fungal Challenge
Abstract
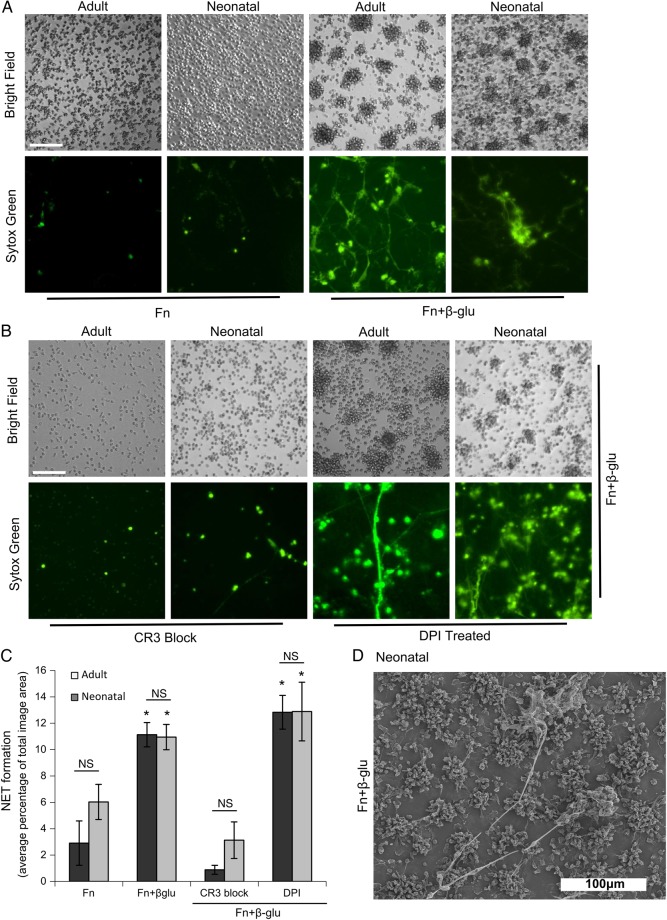
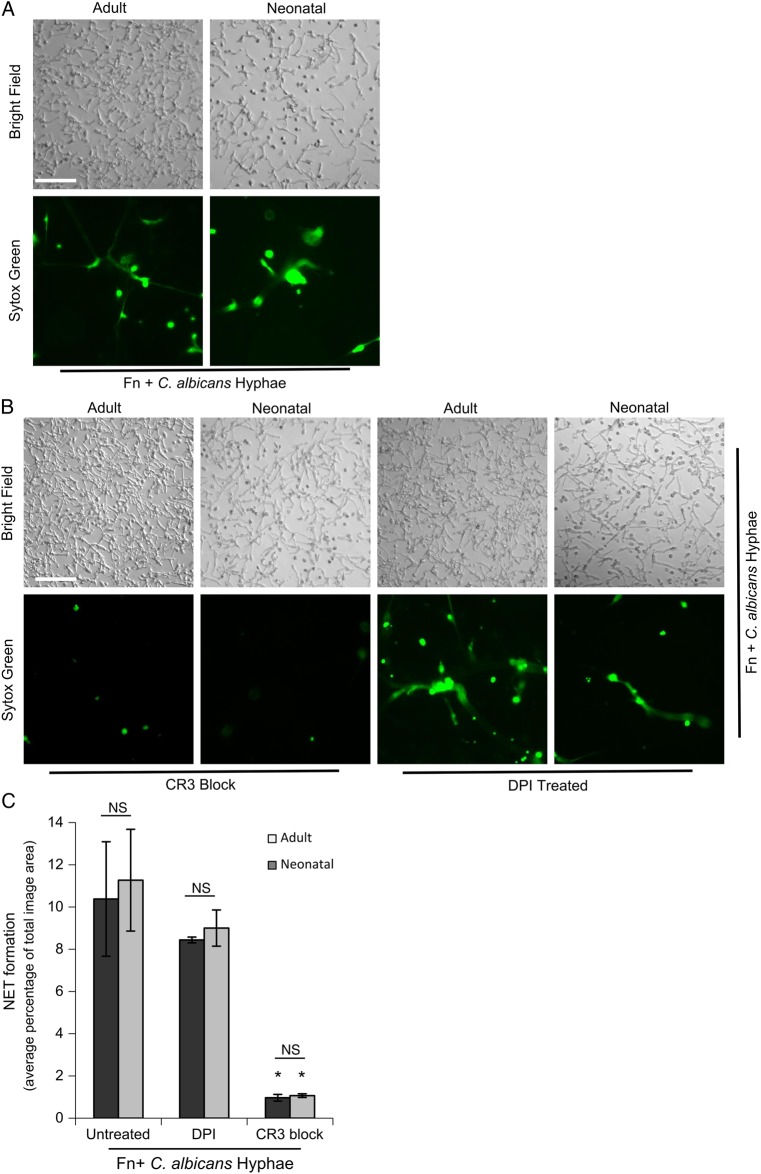
Release of neutrophil extracellular traps (NETs) is a significant antimicrobial host defense mechanism in adults. In neonates, fungal sepsis is a frequent cause of morbidity and mortality and may be a consequence of inadequate neutrophil defense functions. Like neutrophils from adult donors, we found that neutrophils from neonates formed robust cellular aggregates and released NETs in response to fungal β-glucan and Candida albicans hyphae when presented with extracellular matrix. Therefore, in response to fungal stimulation, neonatal neutrophils are capable of NETosis. Neonate susceptibility to fungal infections may not be due to an inability of their neutrophils to produce NETs.
Keywords: NETs; fungi; glucan; neonates; neutrophils.
Published by Oxford University Press for the Infectious Diseases Society of America 2015. This work is written by (a) US Government employee(s) and is in the public domain in the US.
Figures
References
-
- Bardoel BW, Kenny EF, Sollberger G, Zychlinsky A. The balancing act of neutrophils. Cell Host Microbe 2014; 15:526–36. - PubMed
-
- Brinkmann V, Reichard U, Goosmann C et al. . Neutrophil extracellular traps kill bacteria. Science 2004; 303:1532–5. - PubMed
-
- Nathan C, Ding A. Nonresolving inflammation. Cell 2010; 140:871–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
