

Zelda overcomes the high intrinsic nucleosome barrier at enhancers during Drosophila zygotic genome activation
- PMID: 26335633
- PMCID: PMC4617966
- DOI: 10.1101/gr.192542.115
Zelda overcomes the high intrinsic nucleosome barrier at enhancers during Drosophila zygotic genome activation
Abstract
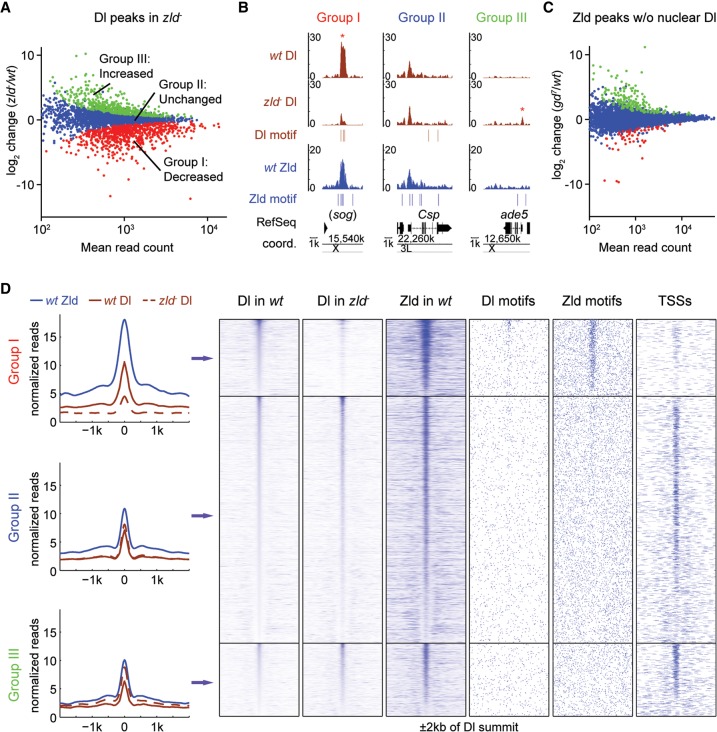
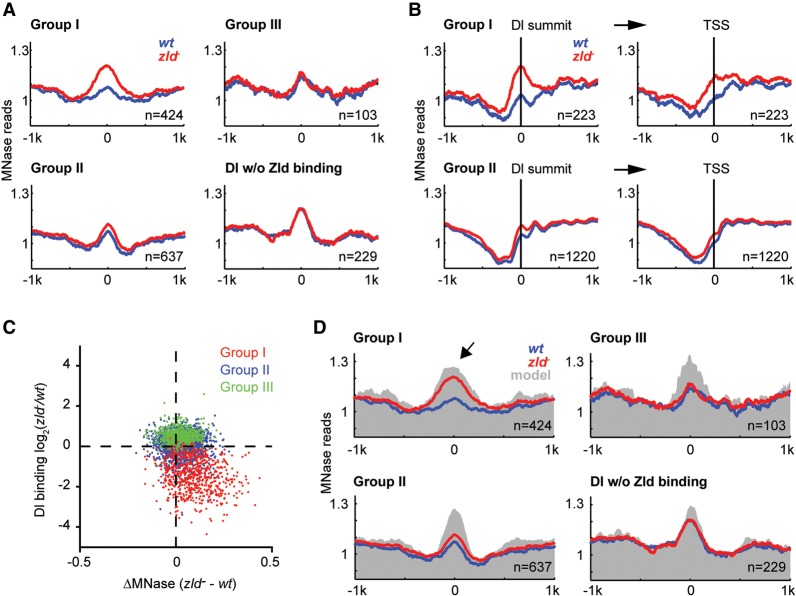
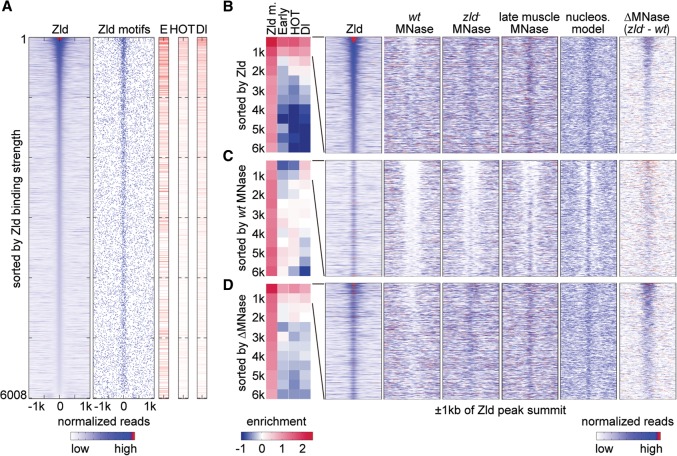
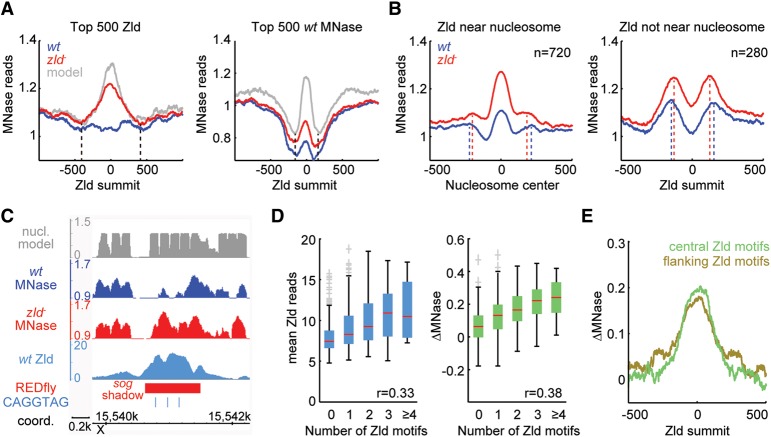
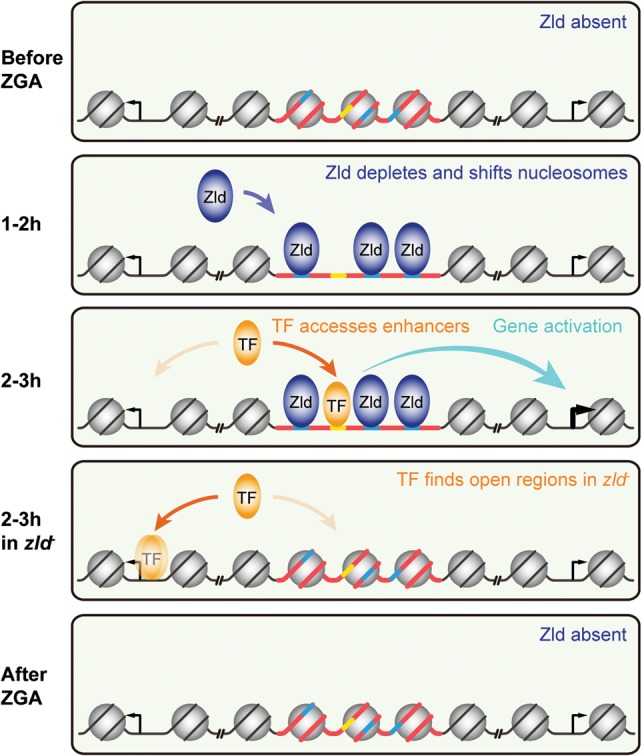
The Drosophila genome activator Vielfaltig (Vfl), also known as Zelda (Zld), is thought to prime enhancers for activation by patterning transcription factors (TFs). Such priming is accompanied by increased chromatin accessibility, but the mechanisms by which this occurs are poorly understood. Here, we analyze the effect of Zld on genome-wide nucleosome occupancy and binding of the patterning TF Dorsal (Dl). Our results show that early enhancers are characterized by an intrinsically high nucleosome barrier. Zld tackles this nucleosome barrier through local depletion of nucleosomes with the effect being dependent on the number and position of Zld motifs. Without Zld, Dl binding decreases at enhancers and redistributes to open regions devoid of enhancer activity. We propose that Zld primes enhancers by lowering the high nucleosome barrier just enough to assist TFs in accessing their binding motifs and promoting spatially controlled enhancer activation if the right patterning TFs are present. We envision that genome activators in general will utilize this mechanism to activate the zygotic genome in a robust and precise manner.
© 2015 Sun et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Arnosti DN, Kulkarni MM. 2005. Transcriptional enhancers: intelligent enhanceosomes or flexible billboards? J Cell Biochem 94: 890–898. - PubMed
-
- Berman BP, Pfeiffer BD, Laverty TR, Salzberg SL, Rubin GM, Eisen MB, Celniker SE. 2004. Computational identification of developmental enhancers: conservation and function of transcription factor binding-site clusters in Drosophila melanogaster and Drosophila pseudoobscura. Genome Biol 5: R61. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous