

Neural Control of Breathing and CO2 Homeostasis
- PMID: 26335642
- PMCID: PMC4559867
- DOI: 10.1016/j.neuron.2015.08.001
Neural Control of Breathing and CO2 Homeostasis
Abstract
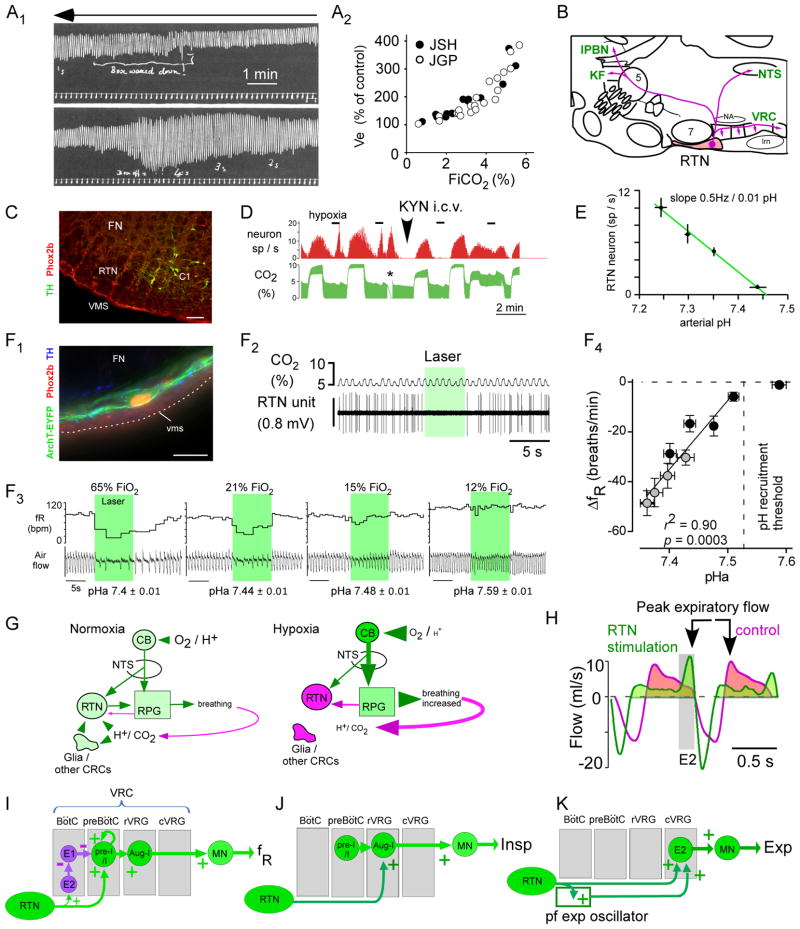
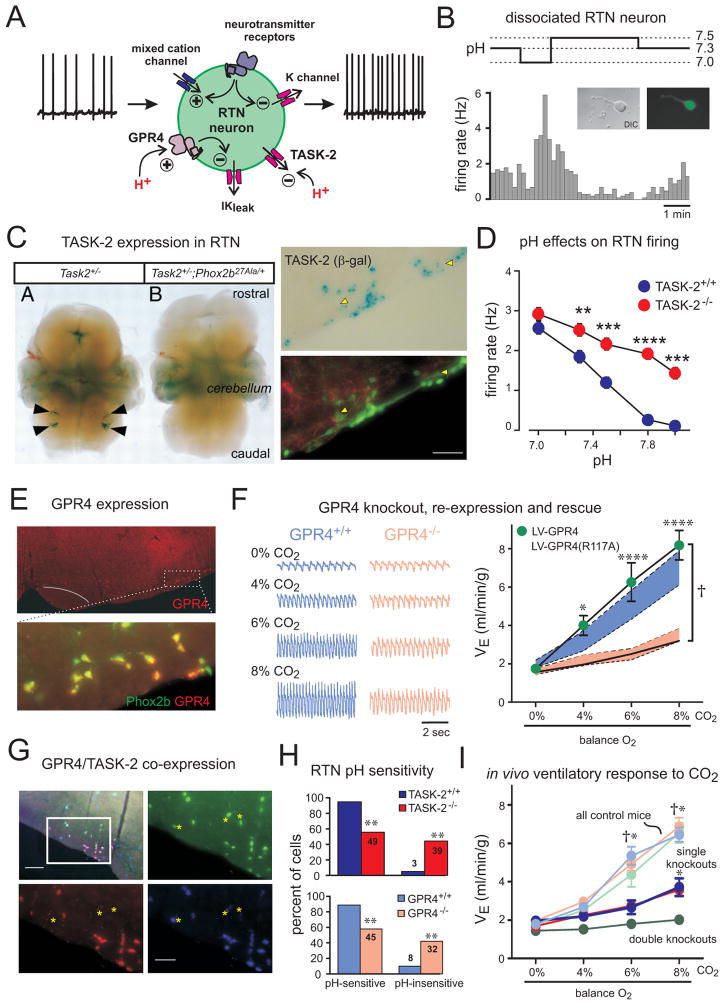
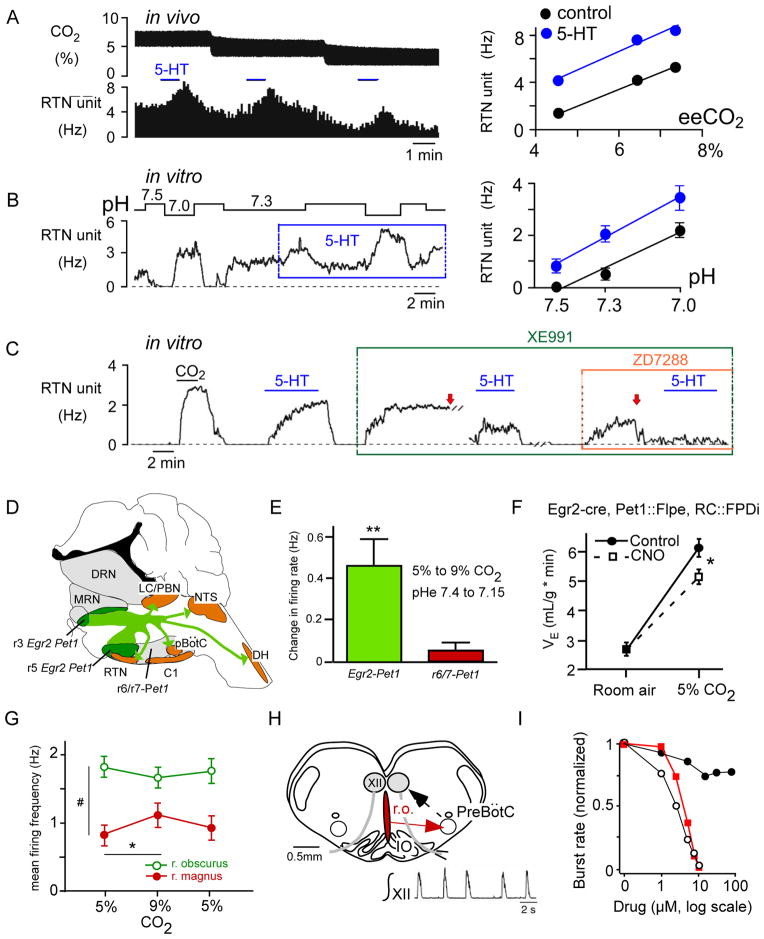
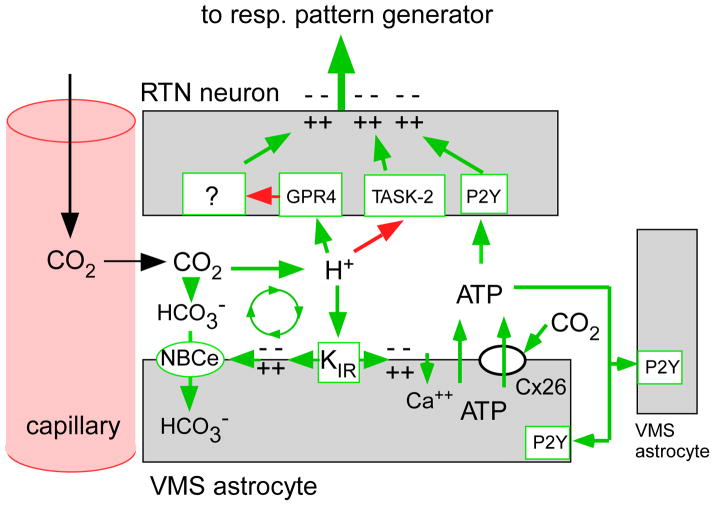
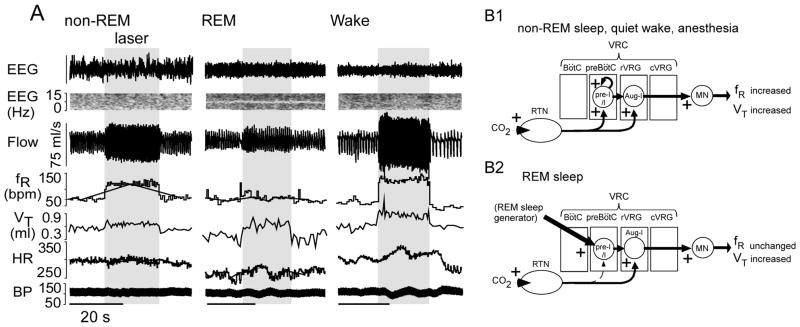
Recent advances have clarified how the brain detects CO2 to regulate breathing (central respiratory chemoreception). These mechanisms are reviewed and their significance is presented in the general context of CO2/pH homeostasis through breathing. At rest, respiratory chemoreflexes initiated at peripheral and central sites mediate rapid stabilization of arterial PCO2 and pH. Specific brainstem neurons (e.g., retrotrapezoid nucleus, RTN; serotonergic) are activated by PCO2 and stimulate breathing. RTN neurons detect CO2 via intrinsic proton receptors (TASK-2, GPR4), synaptic input from peripheral chemoreceptors and signals from astrocytes. Respiratory chemoreflexes are arousal state dependent whereas chemoreceptor stimulation produces arousal. When abnormal, these interactions lead to sleep-disordered breathing. During exercise, central command and reflexes from exercising muscles produce the breathing stimulation required to maintain arterial PCO2 and pH despite elevated metabolic activity. The neural circuits underlying central command and muscle afferent control of breathing remain elusive and represent a fertile area for future investigation.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Amiel J, Laudier B, Attie-Bitach T, Trang H, de PL, Gener B, Trochet D, Etchevers H, Ray P, Simonneau M, Vekemans M, Munnich A, Gaultier C, Lyonnet S. Polyalanine expansion and frameshift mutations of the paired-like homeobox gene PHOX2B in congenital central hypoventilation syndrome. Nat Genet. 2003;33:459–461. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
