

Competing Lipid-Protein and Protein-Protein Interactions Determine Clustering and Gating Patterns in the Potassium Channel from Streptomyces lividans (KcsA)
- PMID: 26336105
- PMCID: PMC4646216
- DOI: 10.1074/jbc.M115.669598
Competing Lipid-Protein and Protein-Protein Interactions Determine Clustering and Gating Patterns in the Potassium Channel from Streptomyces lividans (KcsA)
Abstract
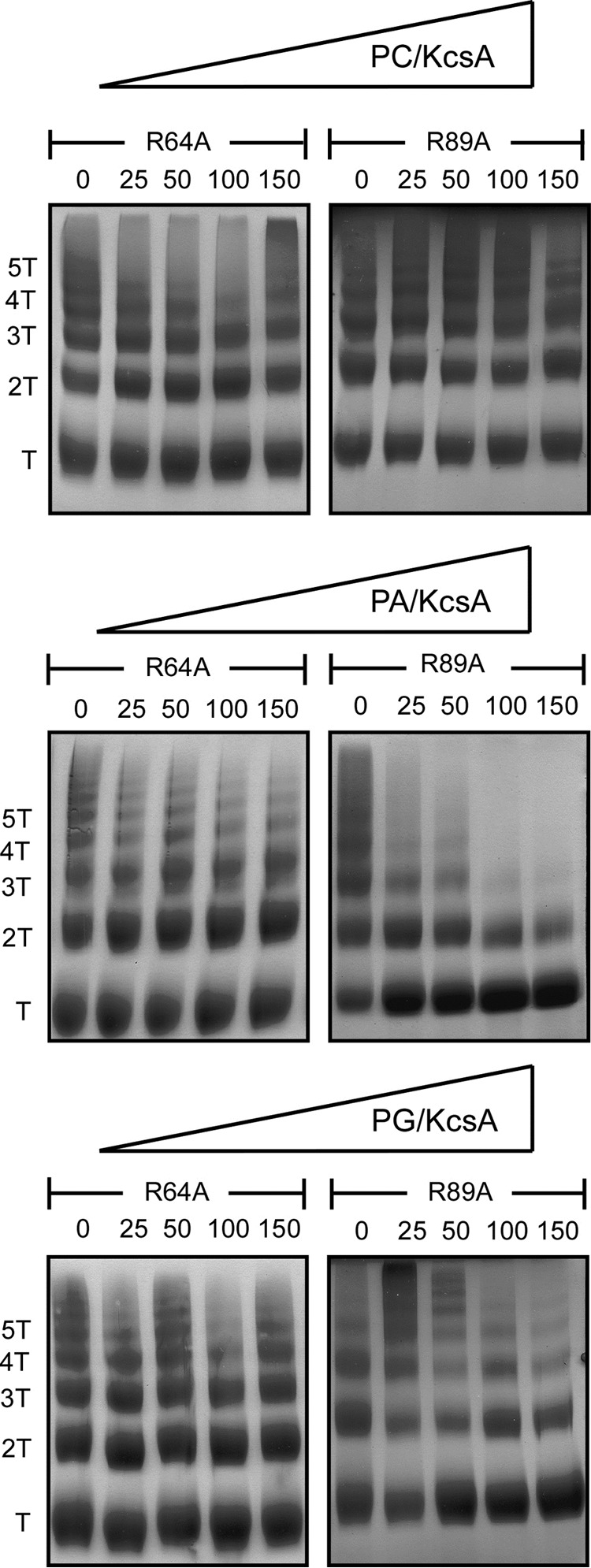
There is increasing evidence to support the notion that membrane proteins, instead of being isolated components floating in a fluid lipid environment, can be assembled into supramolecular complexes that take part in a variety of cooperative cellular functions. The interplay between lipid-protein and protein-protein interactions is expected to be a determinant factor in the assembly and dynamics of such membrane complexes. Here we report on a role of anionic phospholipids in determining the extent of clustering of KcsA, a model potassium channel. Assembly/disassembly of channel clusters occurs, at least partly, as a consequence of competing lipid-protein and protein-protein interactions at nonannular lipid binding sites on the channel surface and brings about profound changes in the gating properties of the channel. Our results suggest that these latter effects of anionic lipids are mediated via the Trp(67)-Glu(71)-Asp(80) inactivation triad within the channel structure and its bearing on the selectivity filter.
Keywords: Ion channel inactivation; Supramolecular assembly of ion channels; coupled channel gating; gating; lipid-protein interaction; membrane protein clusters; membrane protein nonannular sites; molecular modeling; potassium channel; protein-protein interaction.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






References
-
- Doyle D. A., Morais Cabral J., Pfuetzner R. A., Kuo A., Gulbis J. M., Cohen S. L., Chait B. T., MacKinnon R. (1998) The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science 280, 69–77 - PubMed
-
- Zhou Y., Morais-Cabral J. H., Kaufman A., MacKinnon R. (2001) Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 A resolution. Nature 414, 43–48 - PubMed
-
- Meuser D., Splitt H., Wagner R., Schrempf H. (1999) Exploring the open pore of the potassium channel from Streptomyces lividans. FEBS Lett. 462, 447–452 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
