

Phenotypic Correlates of HIV-1 Macrophage Tropism
- PMID: 26339058
- PMCID: PMC4645658
- DOI: 10.1128/JVI.00946-15
Phenotypic Correlates of HIV-1 Macrophage Tropism
Abstract
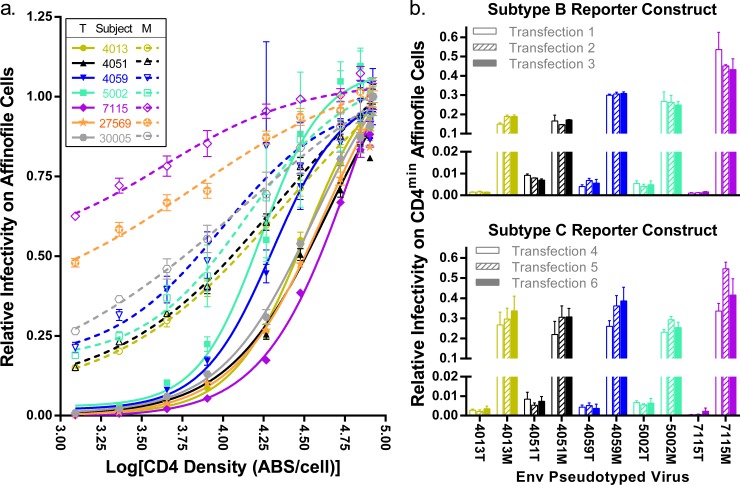
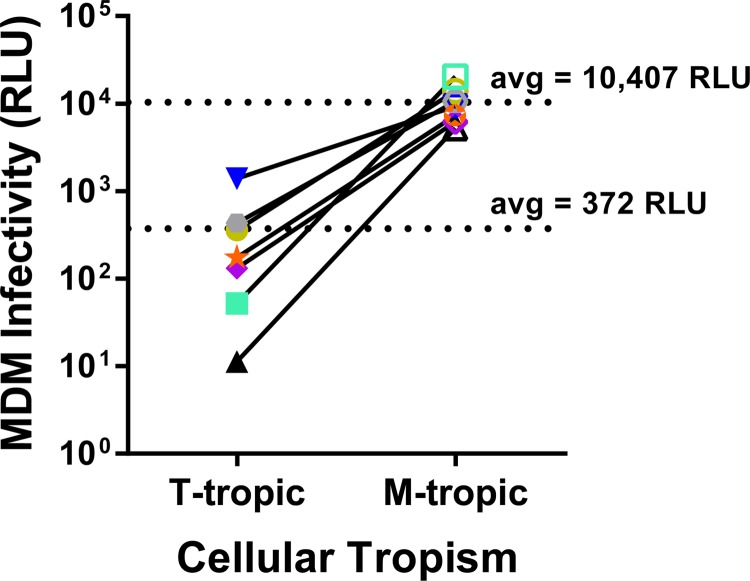
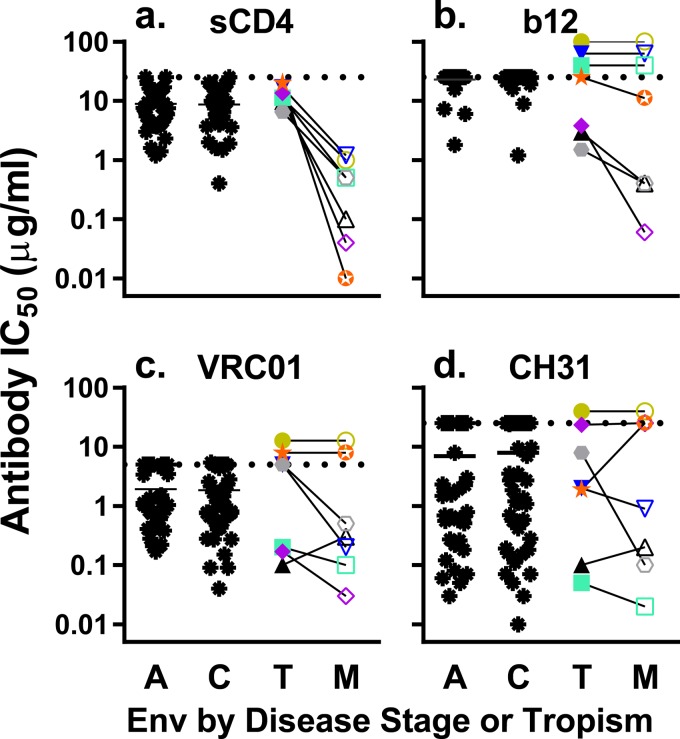
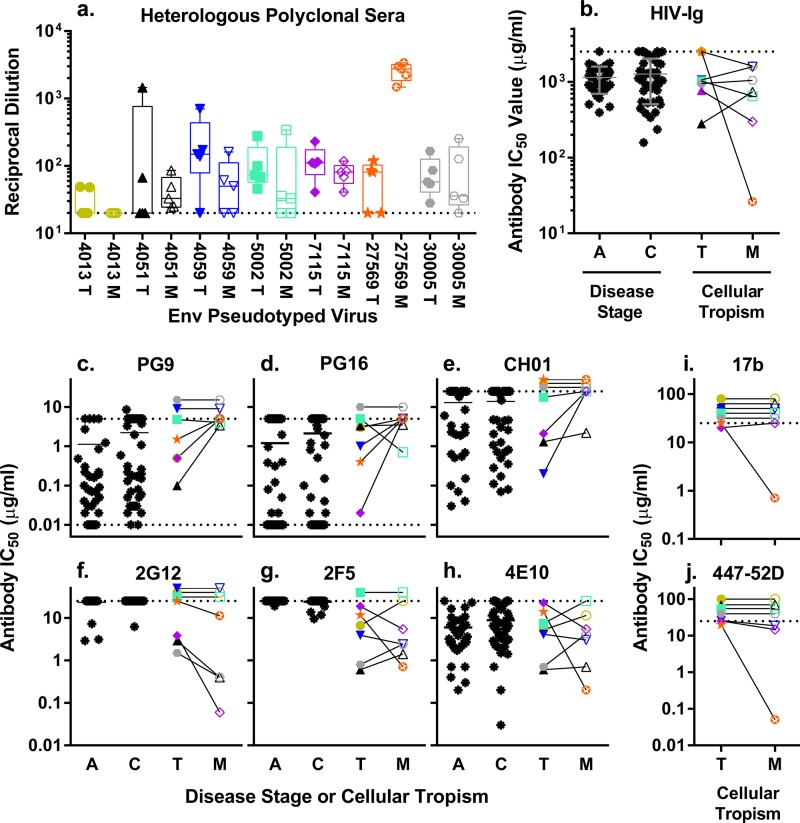
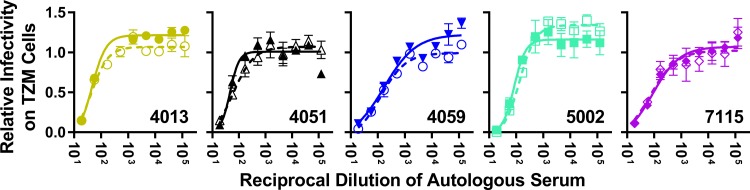
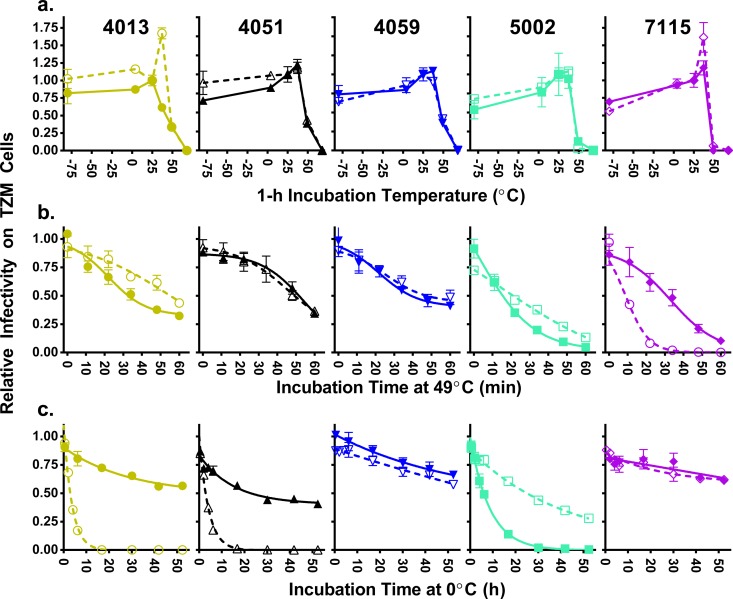
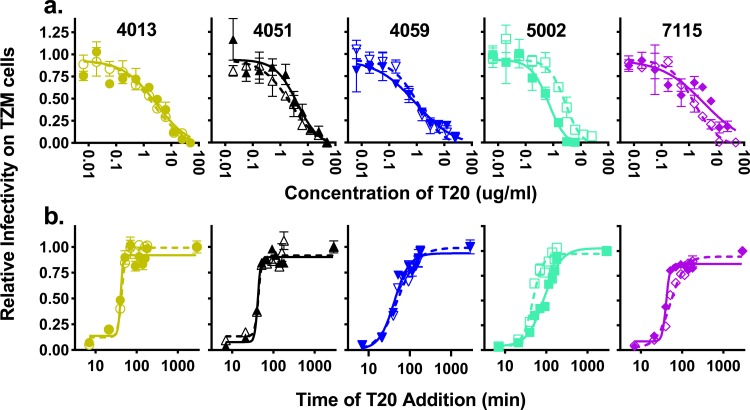
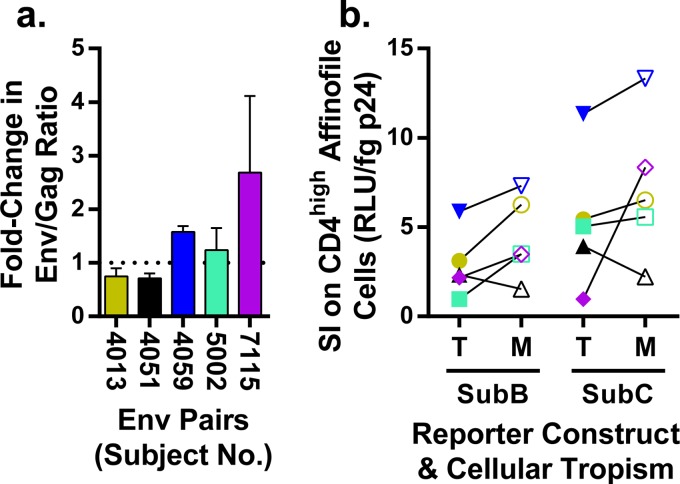
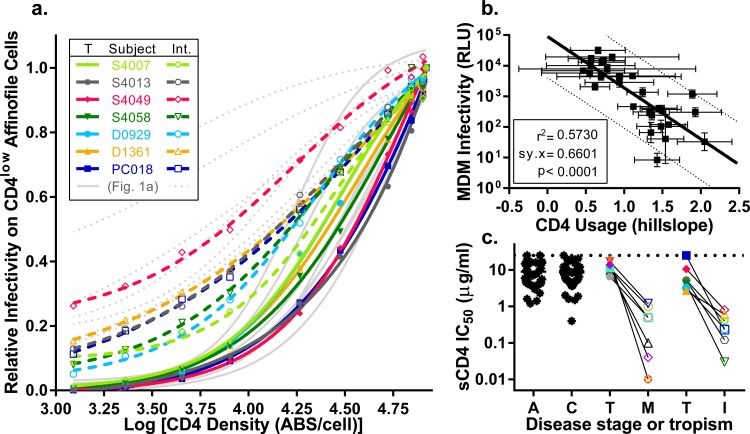
HIV-1 is typically CCR5 using (R5) and T cell tropic (T-tropic), targeting memory CD4(+) T cells throughout acute and chronic infections. However, viruses can expand into alternative cells types. Macrophage-tropic (M-tropic) HIV-1 variants have evolved to infect macrophages, which have only low levels of surface CD4. Most M-tropic variants have been isolated from the central nervous system during late-stage chronic infection. We used the HIV-1 env genes of well-defined, subject-matched M-tropic and T-tropic viruses to characterize the phenotypic features of the M-tropic Env protein. We found that, compared to T-tropic viruses, M-tropic viruses infect monocyte-derived macrophages (MDMs) on average 28-fold more efficiently, use low-density CD4 more efficiently, have increased sensitivity to soluble CD4 (sCD4), and show trends toward sensitivity to some CD4 binding site antibodies but no difference in sensitivity to antibodies targeting the CD4-bound conformation. M-tropic viruses also displayed a trend toward resistance to neutralization by monoclonal antibodies targeting the V1/V2 region of Env, suggesting subtle changes in Env protein conformation. The paired M- and T-tropic viruses did not differ in autologous serum neutralization, temperature sensitivity, entry kinetics, intrinsic infectivity, or Env protein incorporation. We also examined viruses with modestly increased CD4 usage. These variants have significant sensitivity to sCD4 and may represent evolutionary intermediates. CD4 usage is strongly correlated with infectivity of MDMs over a wide range of CD4 entry phenotypes. These data suggest that emergence of M-tropic HIV-1 includes multiple steps in which a phenotype of increased sensitivity to sCD4 and enhanced CD4 usage accompany subtle changes in Env conformation.
Importance: HIV-1 typically replicates in CD4(+) T cells. However, HIV-1 can evolve to infect macrophages, especially within the brain. Understanding how CCR5-using macrophage-tropic viruses evolve and differ from CCR5-using T cell-tropic viruses may provide insights into viral evolution and pathogenesis within the central nervous system. We characterized the HIV-1 env viral entry gene from subject-matched macrophage-tropic and T cell-tropic viruses to identify entry features of macrophage-tropic viruses. We observed several differences between T cell-tropic and macrophage-tropic Env proteins, including functional differences with host CD4 receptor engagement and possible changes in the CD4 binding site and V1/V2 region. We also identified viruses with phenotypes between that of "true" macrophage-tropic and T cell-tropic viruses, which may represent evolutionary intermediates in a multistep process to macrophage tropism.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Isaacman-Beck J, Hermann EA, Yi Y, Ratcliffe SJ, Mulenga J, Allen S, Hunter E, Derdeyn CA, Collman RG. 2009. Heterosexual transmission of human immunodeficiency virus type 1 subtype C: macrophage tropism, alternative coreceptor use, and the molecular anatomy of CCR5 utilization. J Virol 83:8208–8220. doi:10.1128/JVI.00296-09. - DOI - PMC - PubMed
-
- Li M, Salazar-Gonzalez JF, Derdeyn CA, Morris L, Williamson C, Robinson JE, Decker JM, Li Y, Salazar MG, Polonis VR, Mlisana K, Karim SA, Hong K, Greene KM, Bilska M, Zhou J, Allen S, Chomba E, Mulenga J, Vwalika C, Gao F, Zhang M, Korber BTM, Hunter E, Hahn BH, Montefiori DC. 2006. Genetic and neutralization properties of subtype C human immunodeficiency virus type 1 molecular env clones from acute and early heterosexually acquired infections in Southern Africa. J Virol 80:11776–11790. doi:10.1128/JVI.01730-06. - DOI - PMC - PubMed
-
- Ping L-H, Joseph SB, Anderson JA, Abrahams M-R, Salazar-Gonzalez JF, Kincer LP, Treurnicht FK, Arney L, Ojeda S, Zhang M, Keys J, Potter EL, Chu H, Moore P, Salazar MG, Iyer S, Jabara C, Kirchherr J, Mapanje C, Ngandu N, Seoighe C, Hoffman I, Gao F, Tang Y, Labranche C, Lee B, Saville A, Vermeulen M, Fiscus S, Morris L, Karim SA, Haynes BF, Shaw GM, Korber BT, Hahn BH, Cohen MS, Montefiori D, Williamson C, Swanstrom R, CAPRISA Acute Infection Study and the Center for HIV-AIDS Vaccine Immunology Consortium. 2013. Comparison of viral Env proteins from acute and chronic infections with subtype C human immunodeficiency virus type 1 identifies differences in glycosylation and CCR5 utilization and suggests a new strategy for immunogen design. J Virol 87:7218–7233. doi:10.1128/JVI.03577-12. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R37 AI44667/AI/NIAID NIH HHS/United States
- U24MH100925/MH/NIMH NIH HHS/United States
- U24 MH100929/MH/NIMH NIH HHS/United States
- U24 MH100925/MH/NIMH NIH HHS/United States
- U24 MH100931/MH/NIMH NIH HHS/United States
- T32 AI007419/AI/NIAID NIH HHS/United States
- U24MH100928/MH/NIMH NIH HHS/United States
- U24 MH100928/MH/NIMH NIH HHS/United States
- U24MH100930/MH/NIMH NIH HHS/United States
- P30 CA016086/CA/NCI NIH HHS/United States
- U24MH100931/MH/NIMH NIH HHS/United States
- T32 AI07419/AI/NIAID NIH HHS/United States
- P30 AI50410/AI/NIAID NIH HHS/United States
- P30 CA16086/CA/NCI NIH HHS/United States
- R37 AI044667/AI/NIAID NIH HHS/United States
- R01 MH101024/MH/NIMH NIH HHS/United States
- U24MH100929/MH/NIMH NIH HHS/United States
- P30 AI027763/AI/NIAID NIH HHS/United States
- Wellcome Trust/United Kingdom
- U24 MH100930/MH/NIMH NIH HHS/United States
- P01 MH094177/MH/NIMH NIH HHS/United States
- HHSN27201100016C/PHS HHS/United States
- R01 NS094067/NS/NINDS NIH HHS/United States
- P30 AI050410/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
