

Ascorbic acid efflux from human brain microvascular pericytes: role of re-uptake
- PMID: 26340060
- PMCID: PMC4624047
- DOI: 10.1002/biof.1227
Ascorbic acid efflux from human brain microvascular pericytes: role of re-uptake
Abstract
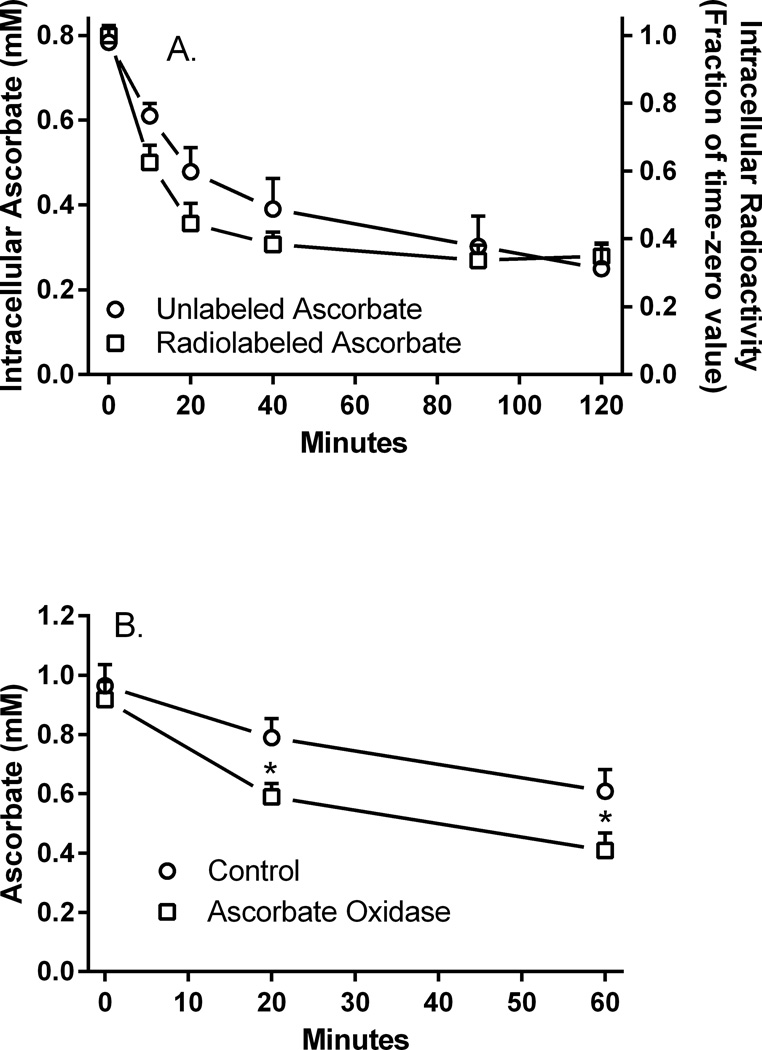
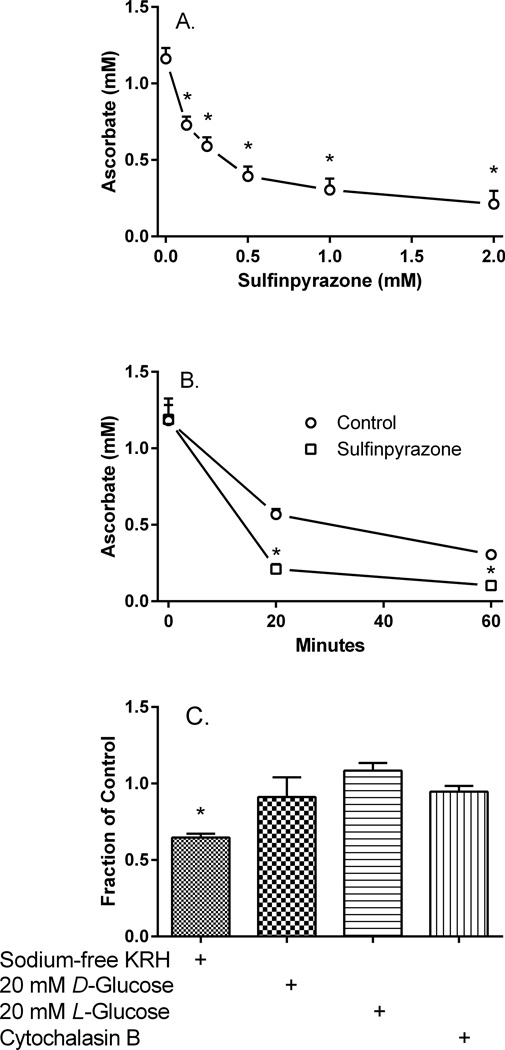
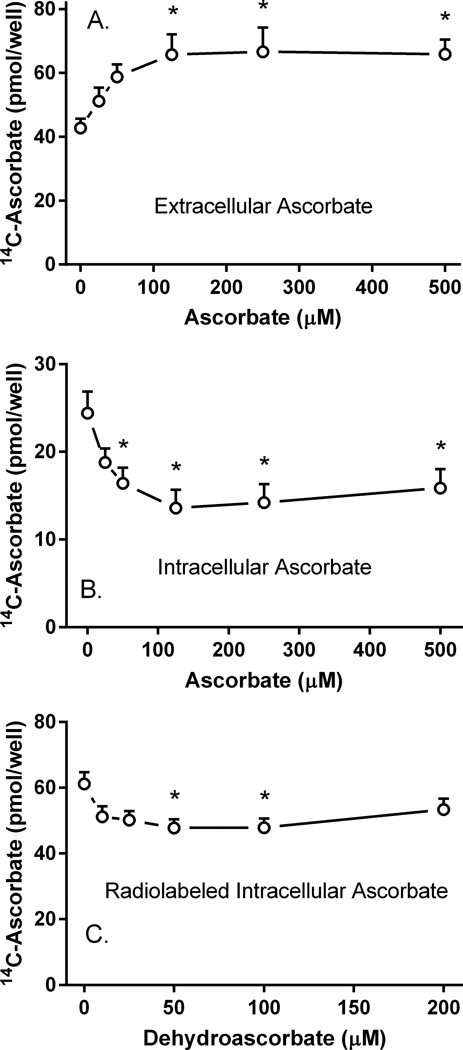
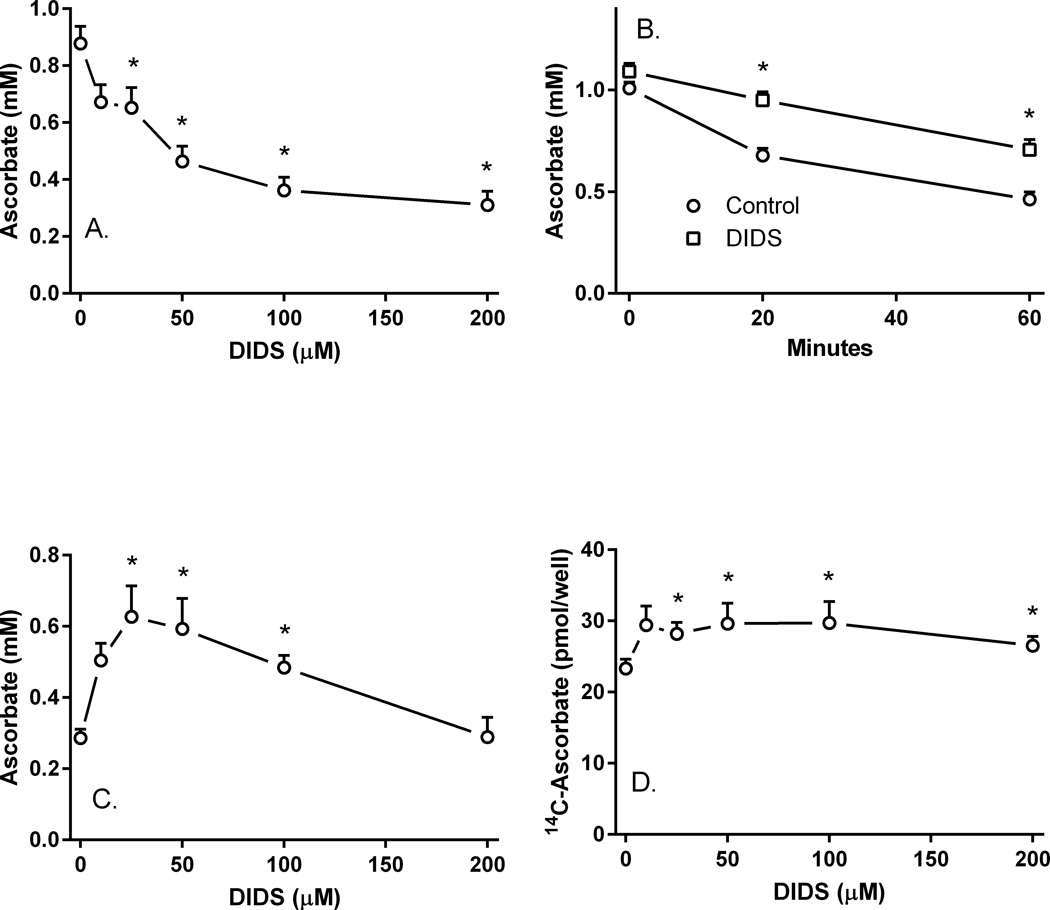
Microvascular pericytes take up ascorbic acid on the ascorbate transporter SVCT2. Intracellular ascorbate then protects the cells against apoptosis induced by culture at diabetic glucose concentrations. To investigate whether pericytes might also provide ascorbate to the underlying endothelial cells, we studied ascorbate efflux from human pericytes. When loaded with ascorbate to intracellular concentrations of 0.8-1.0 mM, almost two-thirds of intracellular ascorbate effluxed from the cells over 2 H. This efflux was opposed by ascorbate re-uptake from the medium, since preventing re-uptake by destroying extracellular ascorbate with ascorbate oxidase increased ascorbate loss even further. Ascorbate re-uptake occurred on the SVCT2, since its blockade by replacing medium sodium with choline, by the SVCT2 inhibitor sulfinpyrazone, or by extracellular ascorbate accelerated ascorbate loss from the cells. This was supported by finding that net efflux of radiolabeled ascorbate was increased by unlabeled extracellular ascorbate with a half-maximal effect in the range of the high affinity Km of the SVCT2. Intracellular ascorbate did not inhibit its efflux. To assess the mechanism of ascorbate efflux, known inhibitors of volume-regulated anion channels (VRACs) were tested. These potently inhibited ascorbate transport into cells on the SVCT2, but not its efflux. An exception was the anion transport inhibitor DIDS, which, despite inhibition of ascorbate uptake, also inhibited net efflux at 25-50 µM. These results suggest that ascorbate efflux from vascular pericytes occurs on a DIDS-inhibitable transporter or channel different from VRACs. Further, ascorbate efflux is opposed by re-uptake of ascorbate on the SVCT2, providing a potential regulatory mechanism.
Keywords: SVCT2; ascorbate efflux; ascorbic acid; pericytes; voltage-regulated anion channels.
© 2015 International Union of Biochemistry and Molecular Biology.
Figures

Similar articles
-
Ascorbic acid transport in brain microvascular pericytes.Biochem Biophys Res Commun. 2015 Mar 6;458(2):262-7. doi: 10.1016/j.bbrc.2015.01.096. Epub 2015 Jan 31. Biochem Biophys Res Commun. 2015. PMID: 25645015 Free PMC article.
-
Mitochondrial ascorbic acid transport is mediated by a low-affinity form of the sodium-coupled ascorbic acid transporter-2.Free Radic Biol Med. 2014 May;70:241-54. doi: 10.1016/j.freeradbiomed.2014.02.021. Epub 2014 Mar 2. Free Radic Biol Med. 2014. PMID: 24594434
-
Sodium-ascorbate cotransport controls intracellular ascorbate concentration in primary astrocyte cultures expressing the SVCT2 transporter.Brain Res. 2000 Oct 27;881(2):144-51. doi: 10.1016/s0006-8993(00)02829-8. Brain Res. 2000. PMID: 11036152
-
The SLC23 family of ascorbate transporters: ensuring that you get and keep your daily dose of vitamin C.Br J Pharmacol. 2011 Dec;164(7):1793-801. doi: 10.1111/j.1476-5381.2011.01350.x. Br J Pharmacol. 2011. PMID: 21418192 Free PMC article. Review.
-
The sodium-dependent ascorbic acid transporter family SLC23.Mol Aspects Med. 2013 Apr-Jun;34(2-3):436-54. doi: 10.1016/j.mam.2012.12.002. Mol Aspects Med. 2013. PMID: 23506882 Review.
Cited by
-
TGF-Beta Modulates the Integrity of the Blood Brain Barrier In Vitro, and Is Associated with Metabolic Alterations in Pericytes.Biomedicines. 2023 Jan 14;11(1):214. doi: 10.3390/biomedicines11010214. Biomedicines. 2023. PMID: 36672722 Free PMC article.
-
The Pharmacokinetics of Vitamin C.Nutrients. 2019 Oct 9;11(10):2412. doi: 10.3390/nu11102412. Nutrients. 2019. PMID: 31601028 Free PMC article. Review.
-
Vitamin C Transporters in Cancer: Current Understanding and Gaps in Knowledge.Front Oncol. 2017 Apr 24;7:74. doi: 10.3389/fonc.2017.00074. eCollection 2017. Front Oncol. 2017. PMID: 28484682 Free PMC article. Review.
-
Role and Molecular Mechanisms of Pericytes in Regulation of Leukocyte Diapedesis in Inflamed Tissues.Mediators Inflamm. 2019 May 7;2019:4123605. doi: 10.1155/2019/4123605. eCollection 2019. Mediators Inflamm. 2019. PMID: 31205449 Free PMC article. Review.
References
-
- Dohgu S, Takata F, Yamauchi A, Nakagawa S, Egawa T, Naito M, Tsuruo T, Sawada Y, Niwa M, Kataoka Y. Brain pericytes contribute to the induction and up-regulation of blood-brain barrier functions through transforming growth factor-beta production. Brain Res. 2005;1038:208–215. - PubMed
-
- Hatherell K, Couraud PO, Romero IA, Weksler B, Pilkington GJ. Development of a three-dimensional, all-human in vitro model of the blood-brain barrier using mono-, co-, and tri-cultivation Transwell models. J Neurosci. Methods. 2011;199:223–229. - PubMed
-
- Wisniewska-Kruk J, Hoeben KA, Vogels IM, Gaillard PJ, Van Noorden CJ, Schlingemann RO, Klaassen I. A novel co-culture model of the blood-retinal barrier based on primary retinal endothelial cells, pericytes and astrocytes. Exp. Eye Res. 2012;96:181–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
