

Negative Regulation of p21Waf1/Cip1 by Human INO80 Chromatin Remodeling Complex Is Implicated in Cell Cycle Phase G2/M Arrest and Abnormal Chromosome Stability
- PMID: 26340092
- PMCID: PMC4560432
- DOI: 10.1371/journal.pone.0137411
Negative Regulation of p21Waf1/Cip1 by Human INO80 Chromatin Remodeling Complex Is Implicated in Cell Cycle Phase G2/M Arrest and Abnormal Chromosome Stability
Abstract
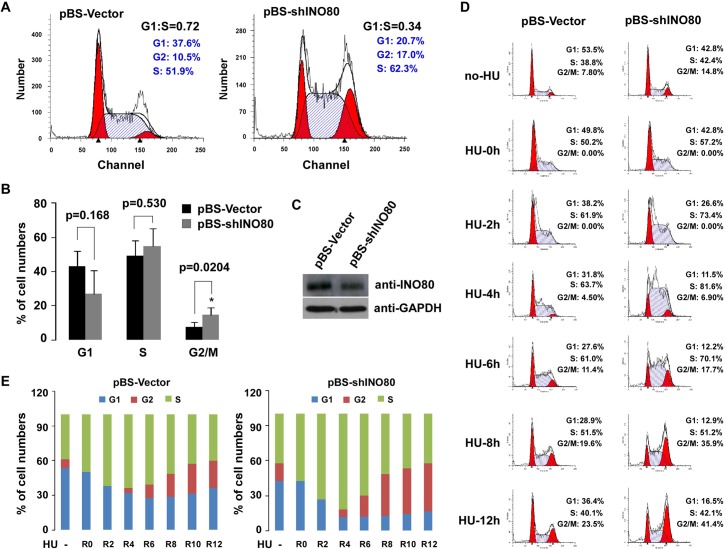
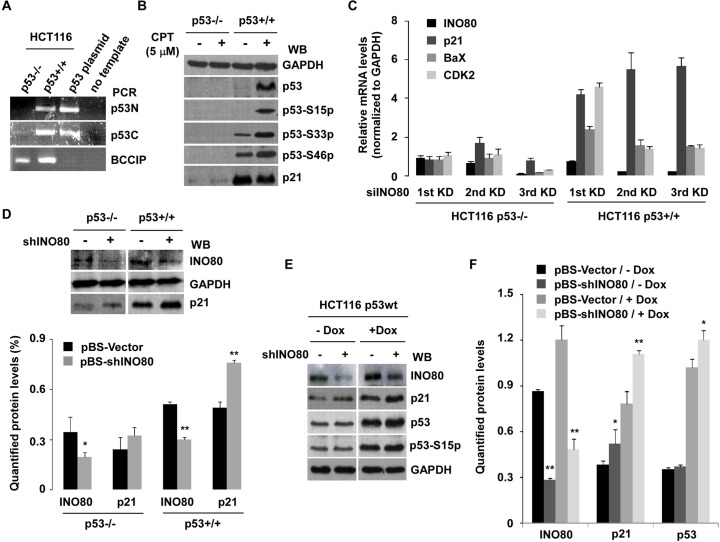
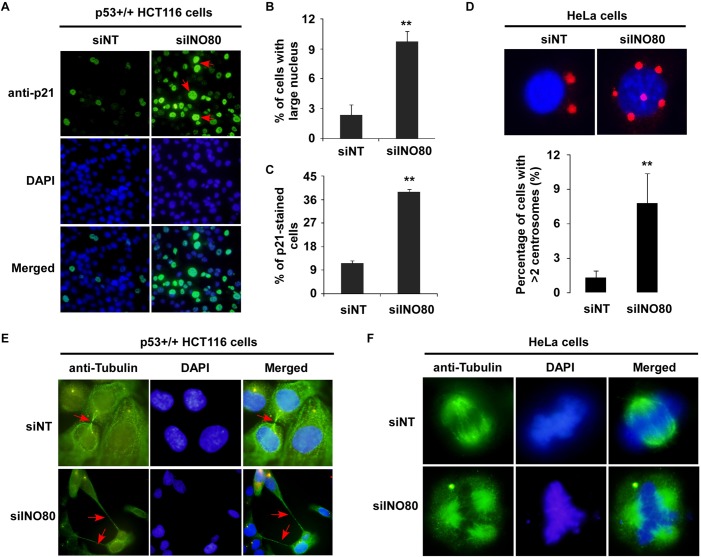
We previously identified an ATP-dependent human Ino80 (INO80) chromatin remodeling complex which shares a set of core subunits with yeast Ino80 complex. Although research evidence has suggested that INO80 complex functions in gene transcription and genome stability, the precise mechanism remains unclear. Herein, based on gene expression profiles from the INO80 complex-knockdown in HeLa cells, we first demonstrate that INO80 complex negatively regulates the p21Waf1/Cip1 (p21) expression in a p53-mediated mechanism. In chromatin immunoprecipitation (ChIP) and a sequential ChIP (Re-ChIP) assays, we determined that the INO80 complex and p53 can bind to the same promoter region of p21 gene (-2.2 kb and -1.0 kb upstream of the p21 promoter region), and p53 is required for the recruitment of the INO80 complex to the p21 promoter. RNAi knockdown strategies of INO80 not only led to prolonged progression of cell cycle phase G2/M to G1, but it also resulted in abnormal chromosome stability. Interestingly, high expression of p21 was observed in most morphologically-changed cells, suggesting that negative regulation of p21 by INO80 complex might be implicated in maintaining the cell cycle process and chromosome stability. Together, our findings will provide a theoretical basis to further elucidate the cellular mechanisms of the INO80 complex.
Conflict of interest statement
Figures




Similar articles
-
YY1/BCCIP Coordinately Regulates P53-Responsive Element (p53RE)-Mediated Transactivation of p21Waf1/Cip1.Int J Mol Sci. 2019 Apr 28;20(9):2095. doi: 10.3390/ijms20092095. Int J Mol Sci. 2019. PMID: 31035388 Free PMC article.
-
The chromatin remodeling protein INO80 contributes to the removal of H2A.Z at the p53-binding site of the p21 gene in response to doxorubicin.FEBS J. 2018 Sep;285(17):3270-3285. doi: 10.1111/febs.14615. Epub 2018 Aug 14. FEBS J. 2018. PMID: 30055111
-
Human INO80/YY1 chromatin remodeling complex transcriptionally regulates the BRCA2- and CDKN1A-interacting protein (BCCIP) in cells.Protein Cell. 2016 Oct;7(10):749-760. doi: 10.1007/s13238-016-0306-1. Epub 2016 Aug 17. Protein Cell. 2016. PMID: 27535137 Free PMC article.
-
The INO80 family of chromatin-remodeling enzymes: regulators of histone variant dynamics.Cold Spring Harb Symp Quant Biol. 2010;75:35-42. doi: 10.1101/sqb.2010.75.063. Epub 2011 Apr 18. Cold Spring Harb Symp Quant Biol. 2010. PMID: 21502417 Review.
-
Rvb1-Rvb2: essential ATP-dependent helicases for critical complexes.Biochem Cell Biol. 2010 Feb;88(1):29-40. doi: 10.1139/o09-122. Biochem Cell Biol. 2010. PMID: 20130677 Review.
Cited by
-
YY1/BCCIP Coordinately Regulates P53-Responsive Element (p53RE)-Mediated Transactivation of p21Waf1/Cip1.Int J Mol Sci. 2019 Apr 28;20(9):2095. doi: 10.3390/ijms20092095. Int J Mol Sci. 2019. PMID: 31035388 Free PMC article.
-
ATP-Dependent Chromatin Remodeler CHD9 Controls the Proliferation of Embryonic Stem Cells in a Cell Culture Condition-Dependent Manner.Biology (Basel). 2020 Nov 27;9(12):428. doi: 10.3390/biology9120428. Biology (Basel). 2020. PMID: 33261017 Free PMC article.
-
Ino80 is required for H2A.Z eviction from hypha-specific promoters and hyphal development of Candida albicans.Mol Microbiol. 2022 Jul;118(1-2):92-104. doi: 10.1111/mmi.14954. Epub 2022 Jun 28. Mol Microbiol. 2022. PMID: 35713098 Free PMC article.
-
INO80 Is Required for the Cell Cycle Control, Survival, and Differentiation of Mouse ESCs by Transcriptional Regulation.Int J Mol Sci. 2022 Dec 6;23(23):15402. doi: 10.3390/ijms232315402. Int J Mol Sci. 2022. PMID: 36499727 Free PMC article.
-
HIRA complex presets transcriptional potential through coordinating depositions of the histone variants H3.3 and H2A.Z on the poised genes in mESCs.Nucleic Acids Res. 2022 Jan 11;50(1):191-206. doi: 10.1093/nar/gkab1221. Nucleic Acids Res. 2022. PMID: 34893908 Free PMC article.
References
-
- Jin J, Cai Y, Yao T, Gottschalk AJ, Florens F, Swanson SK, et al. A mammalian chromatin remodeling complex with similarities to the yeast INO80 complex. J Biol Chem.2005; 280: 41207–12. - PubMed
-
- Cai Y, Jin J, Yao T, Gottschalk AJ, Swanson SK, Wu S, et al. YY1 functions with INO80 to activate transcription. Nat Struc Mol Biol. 2007; 14: 872–4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
