

Novel identification of the free fatty acid receptor FFAR1 that promotes contraction in airway smooth muscle
- PMID: 26342087
- PMCID: PMC4628981
- DOI: 10.1152/ajplung.00041.2015
Novel identification of the free fatty acid receptor FFAR1 that promotes contraction in airway smooth muscle
Abstract
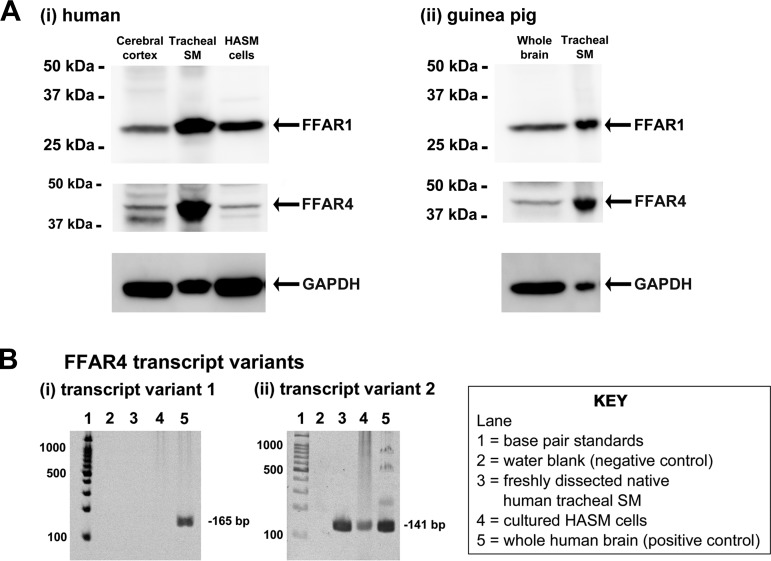
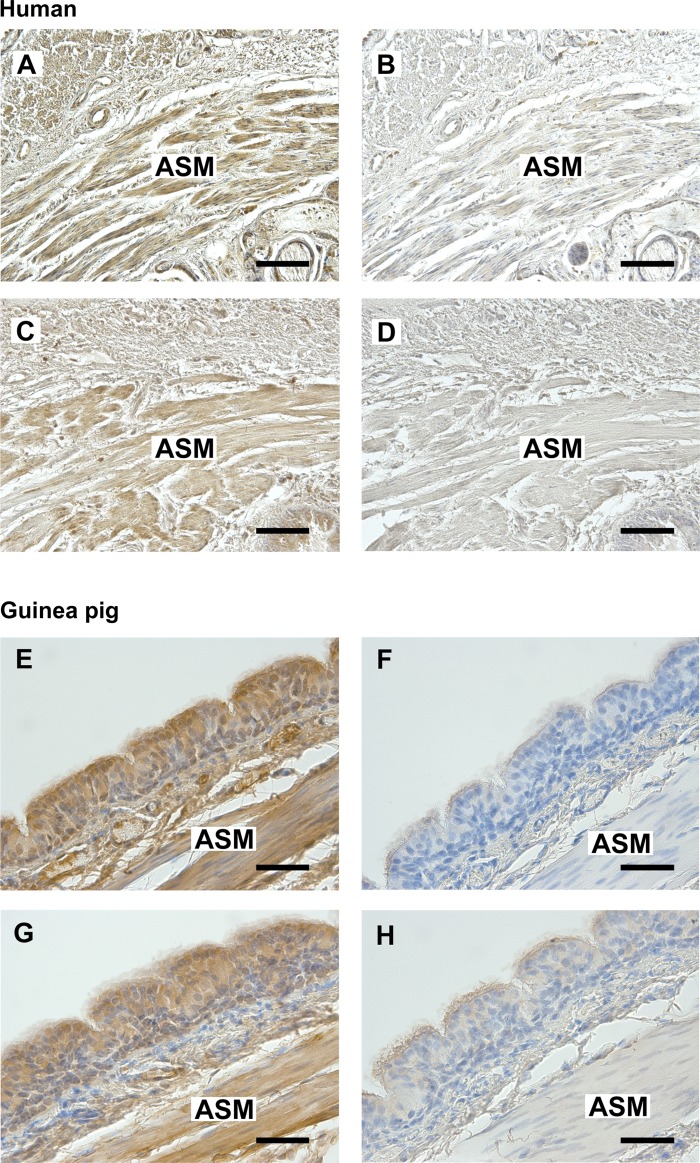
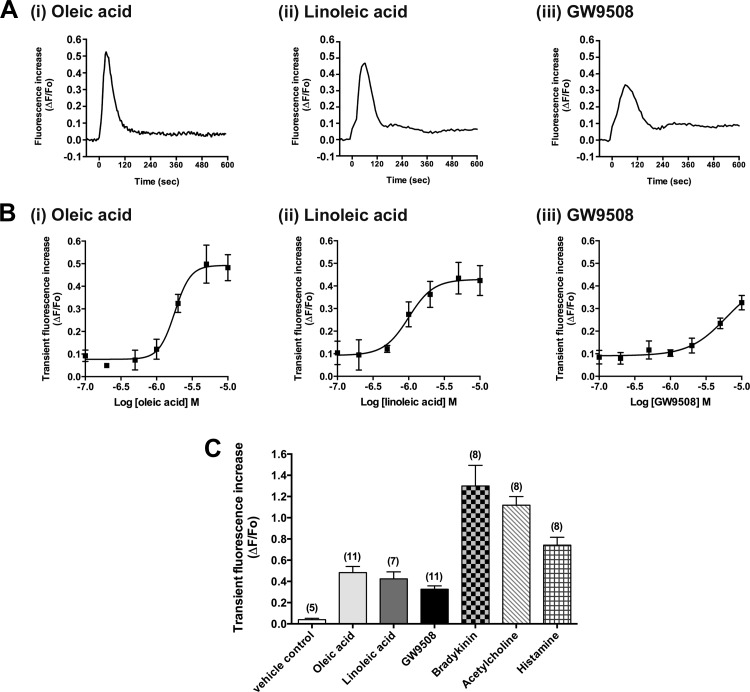
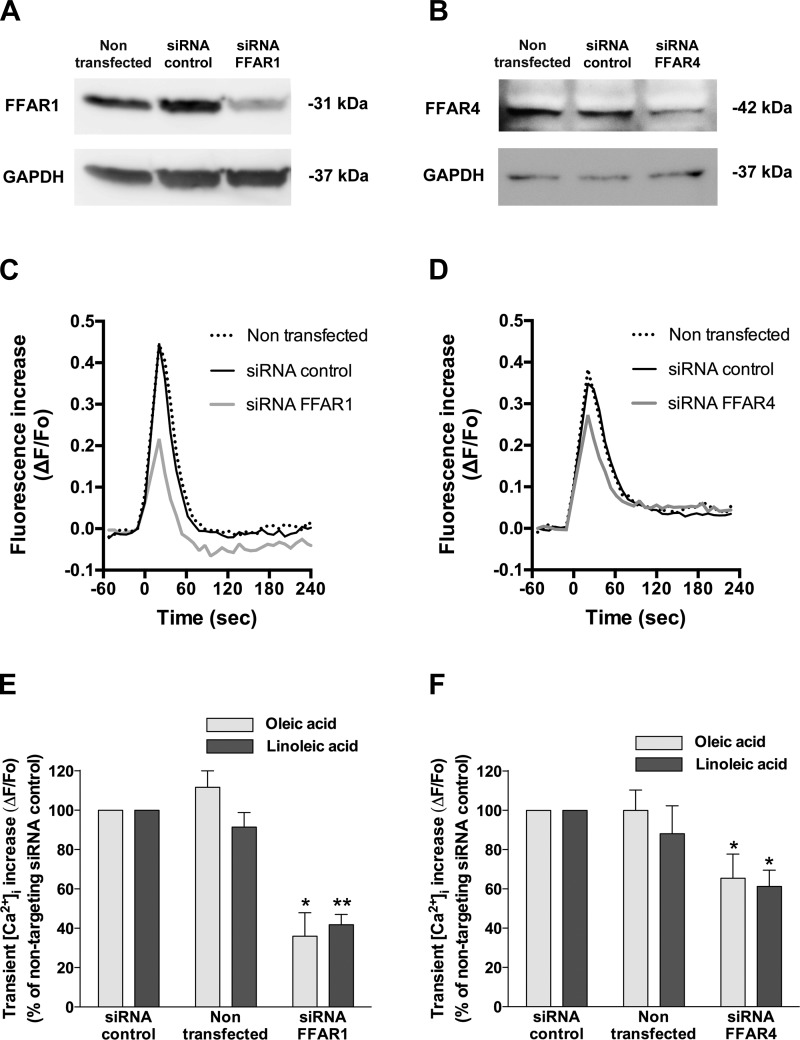
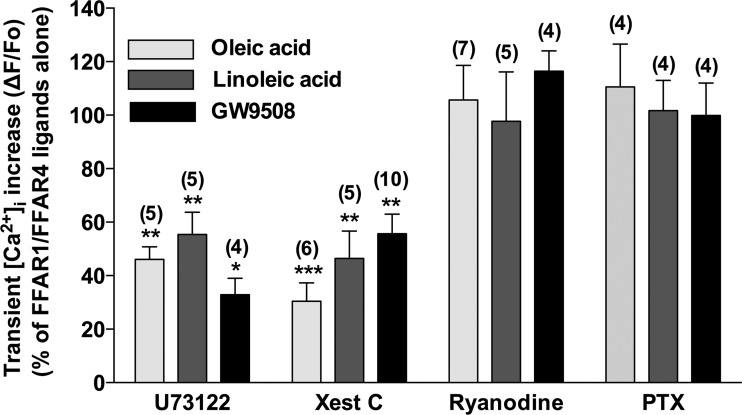
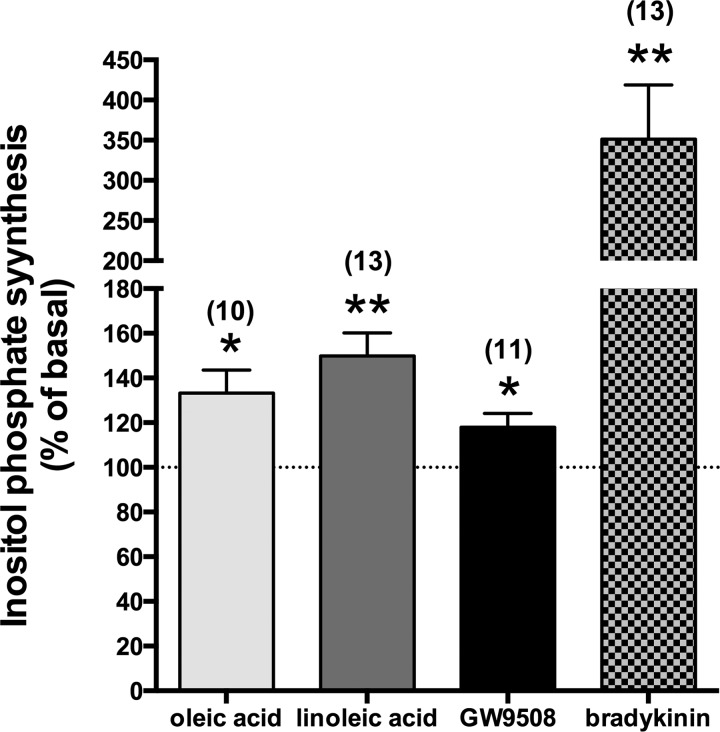
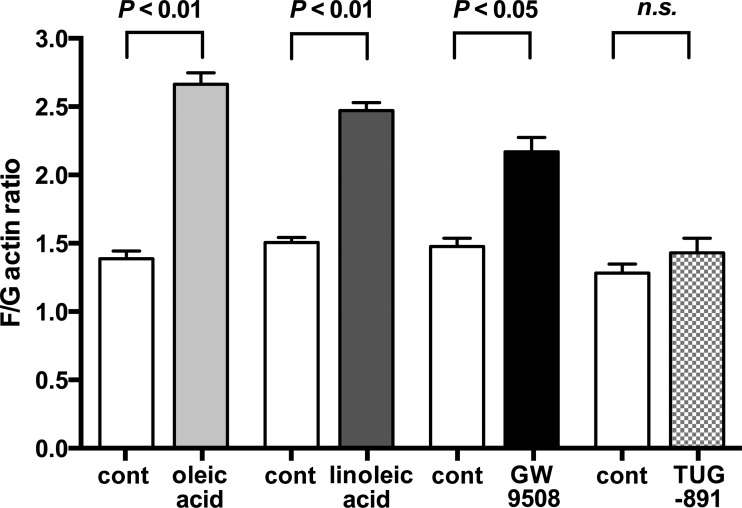
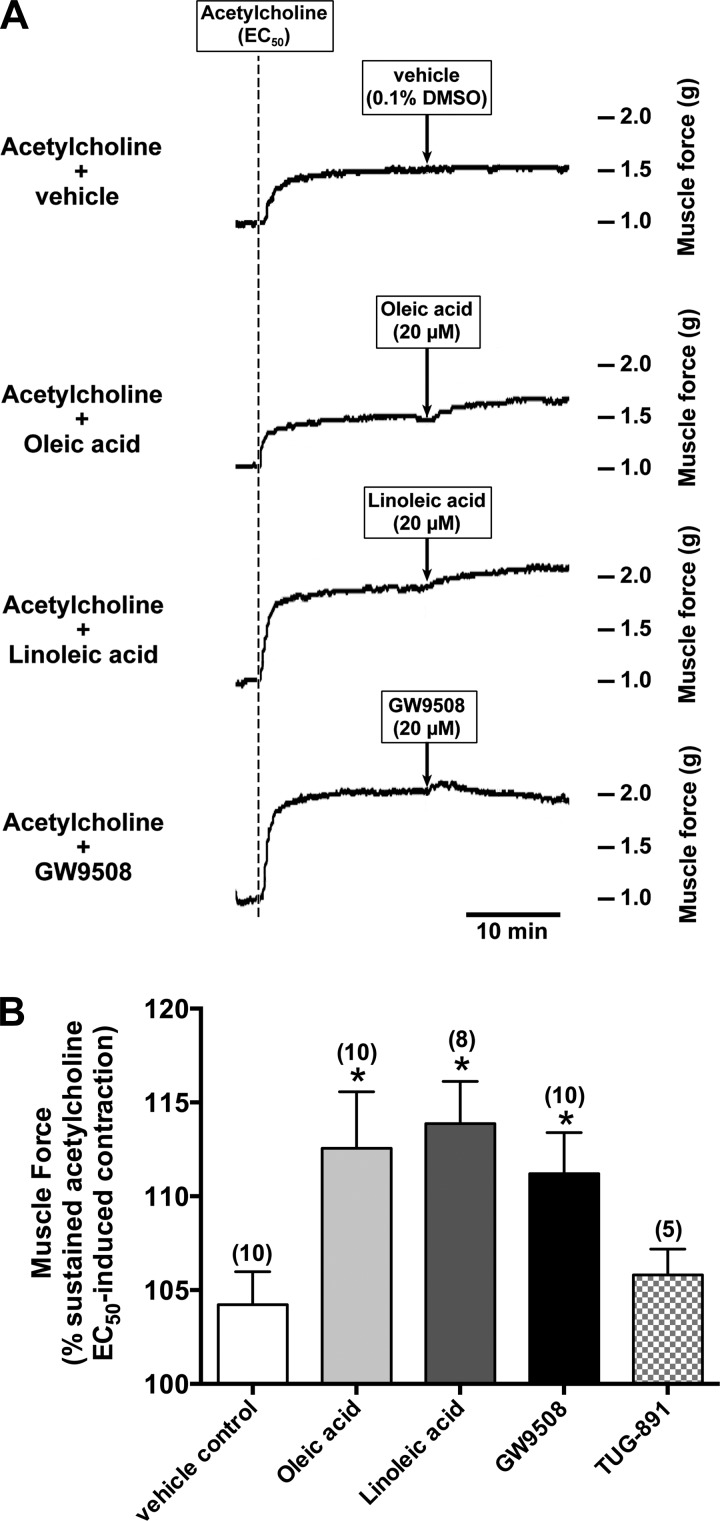
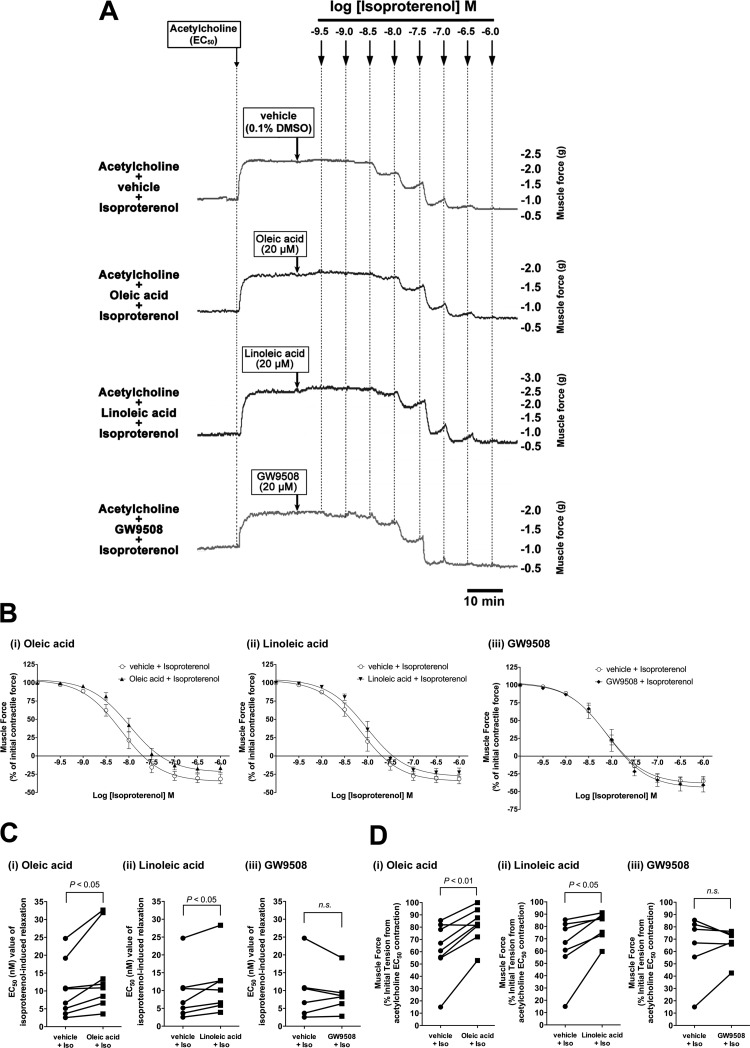
Obesity is one of the major risk factors for asthma. Previous studies have demonstrated that free fatty acid levels are elevated in the plasma of obese individuals. Medium- and long-chain free fatty acids act as endogenous ligands for the free fatty acid receptors FFAR1/GPR40 and FFAR4/GPR120, which couple to Gq proteins. We investigated whether FFAR1 and FFAR4 are expressed on airway smooth muscle and whether they activate Gq-coupled signaling and modulate airway smooth muscle tone. We detected the protein expression of FFAR1 and FFAR4 in freshly dissected native human and guinea pig airway smooth muscle and cultured human airway smooth muscle (HASM) cells by immunoblotting and immunohistochemistry. The long-chain free fatty acids (oleic acid and linoleic acid) and GW9508 (FFAR1/FFAR4 dual agonist) dose-dependently stimulated transient intracellular Ca(2+) concentration ([Ca(2+)]i) increases and inositol phosphate synthesis in HASM cells. Downregulation of FFAR1 or FFAR4 in HASM cells by small interfering RNA led to a significant inhibition of the long-chain free fatty acids-induced transient [Ca(2+)]i increases. Oleic acid, linoleic acid, or GW9508 stimulated stress fiber formation in HASM cells, potentiated acetylcholine-contracted guinea pig tracheal rings, and attenuated the relaxant effect of isoproterenol after an acetylcholine-induced contraction. In contrast, TUG-891 (FFAR4 agonist) did not induce the stress fiber formation or potentiate acetylcholine-induced contraction. These results suggest that FFAR1 is the functionally dominant free fatty acid receptor in both human and guinea pig airway smooth muscle. The free fatty acid sensors expressed on airway smooth muscle could be an important modulator of airway smooth muscle tone.
Keywords: FFAR1; FFAR4; Gq-coupled receptor; airway smooth muscle; free fatty acid.
Copyright © 2015 the American Physiological Society.
Figures
References
-
- Bjorntorp P, Bergman H, Varnauskas E. Plasma free fatty acid turnover rate in obesity. Acta Med Scand 185: 351–356, 1969. - PubMed
-
- Briscoe CP, Peat AJ, McKeown SC, Corbett DF, Goetz AS, Littleton TR, McCoy DC, Kenakin TP, Andrews JL, Ammala C, Fornwald JA, Ignar DM, Jenkinson S. Pharmacological regulation of insulin secretion in MIN6 cells through the fatty acid receptor GPR40: identification of agonist and antagonist small molecules. Br J Pharmacol 148: 619–628, 2006. - PMC - PubMed
-
- Briscoe CP, Tadayyon M, Andrews JL, Benson WG, Chambers JK, Eilert MM, Ellis C, Elshourbagy NA, Goetz AS, Minnick DT, Murdock PR, Sauls HR Jr, Shabon U, Spinage LD, Strum JC, Szekeres PG, Tan KB, Way JM, Ignar DM, Wilson S, Muir AI. The orphan G protein-coupled receptor GPR40 is activated by medium and long chain fatty acids. J Biol Chem 278: 11303–11311, 2003. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous