

Cyclooxygenase-Dependent Tumor Growth through Evasion of Immunity
- PMID: 26343581
- PMCID: PMC4597191
- DOI: 10.1016/j.cell.2015.08.015
Cyclooxygenase-Dependent Tumor Growth through Evasion of Immunity
Abstract
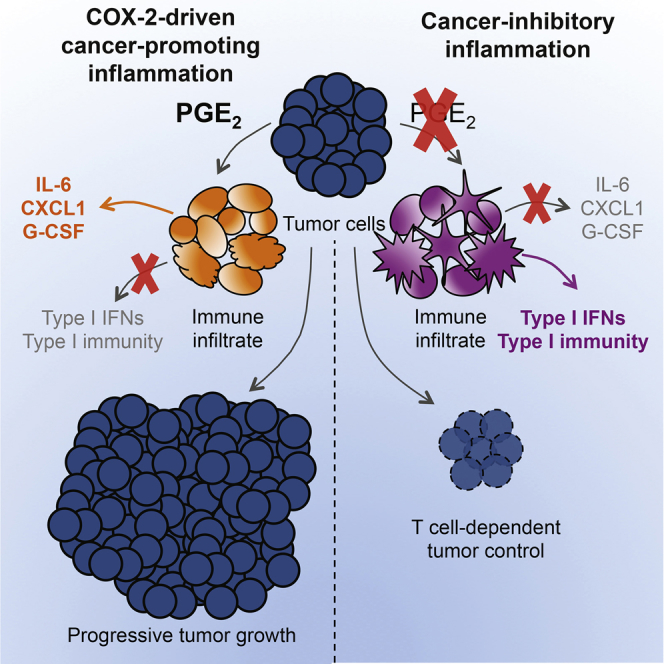
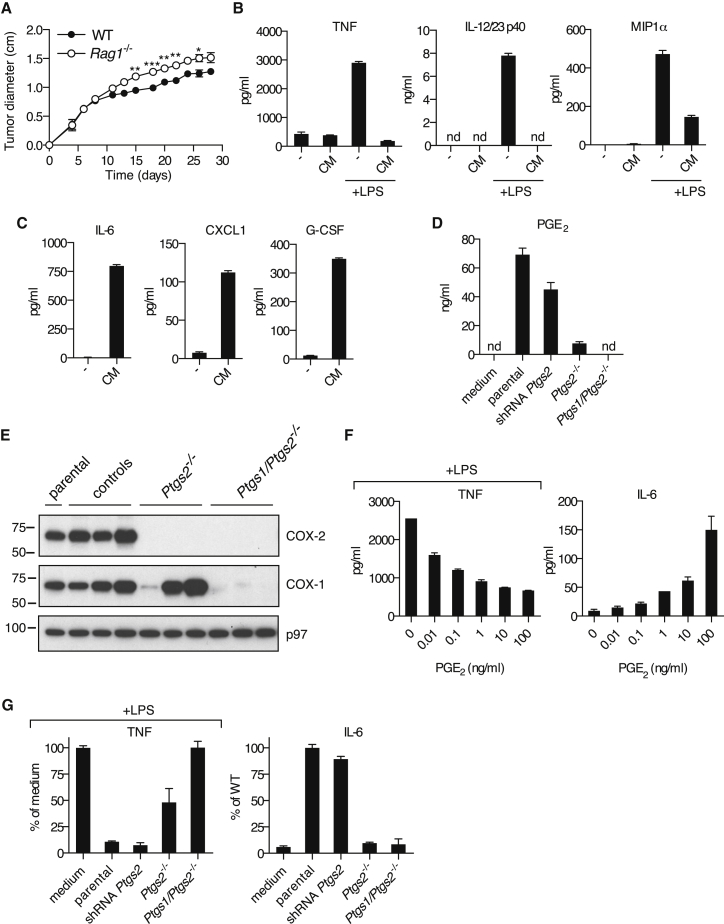
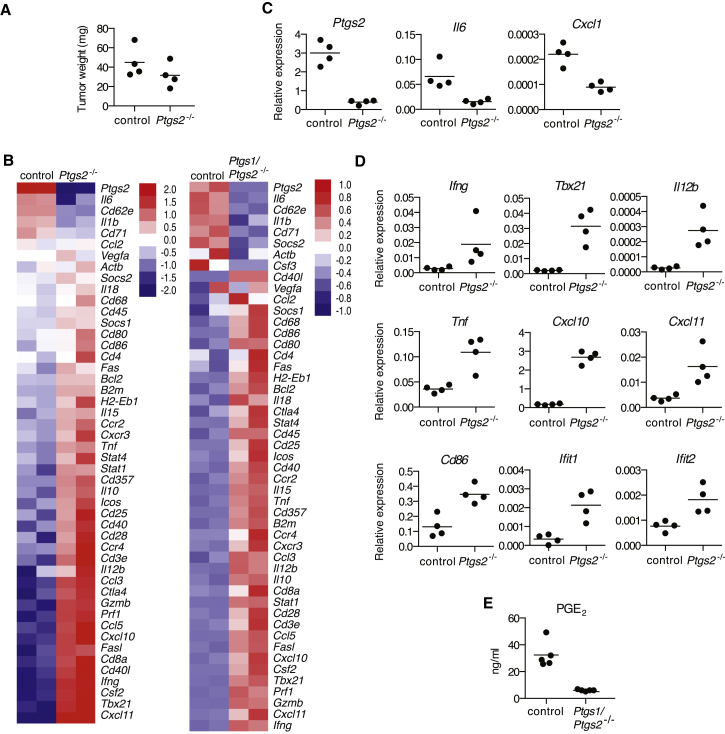
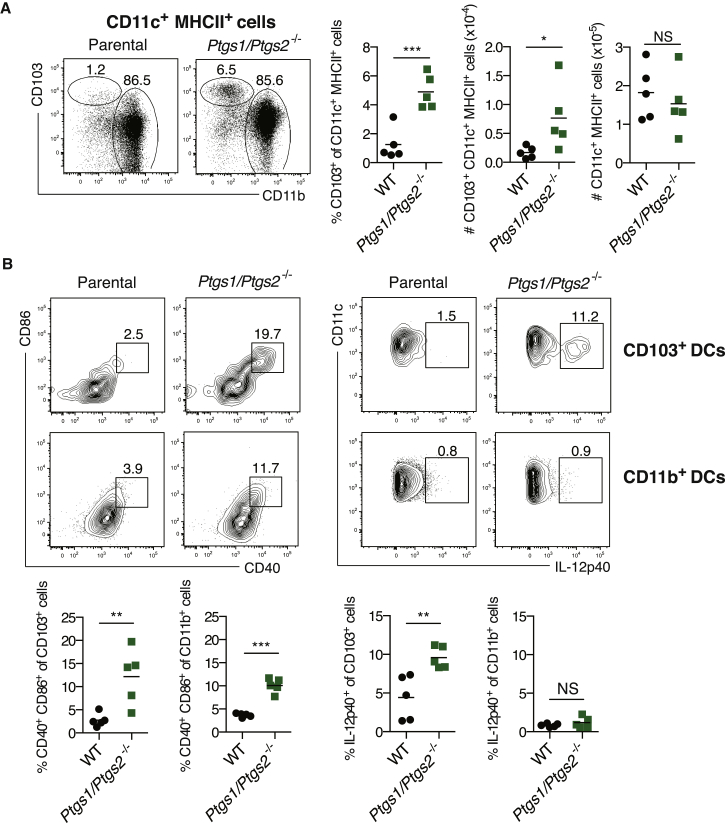
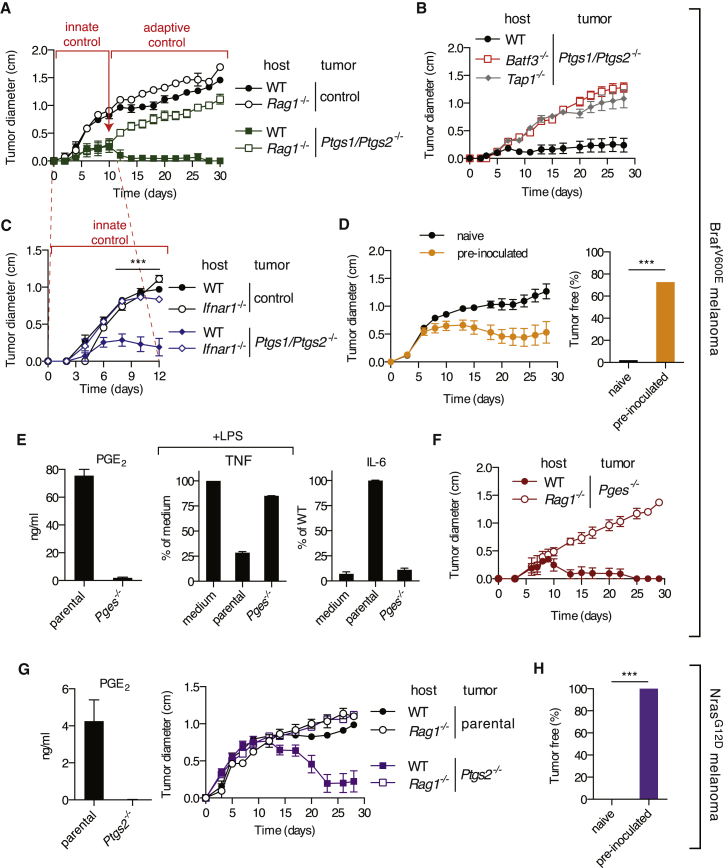
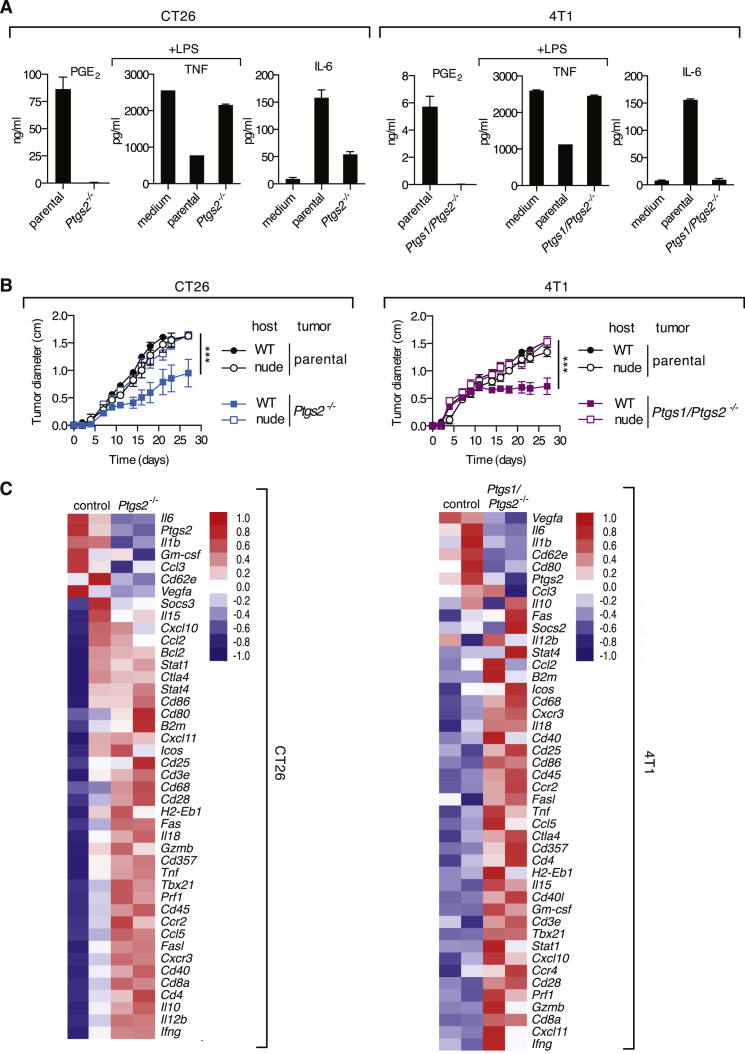
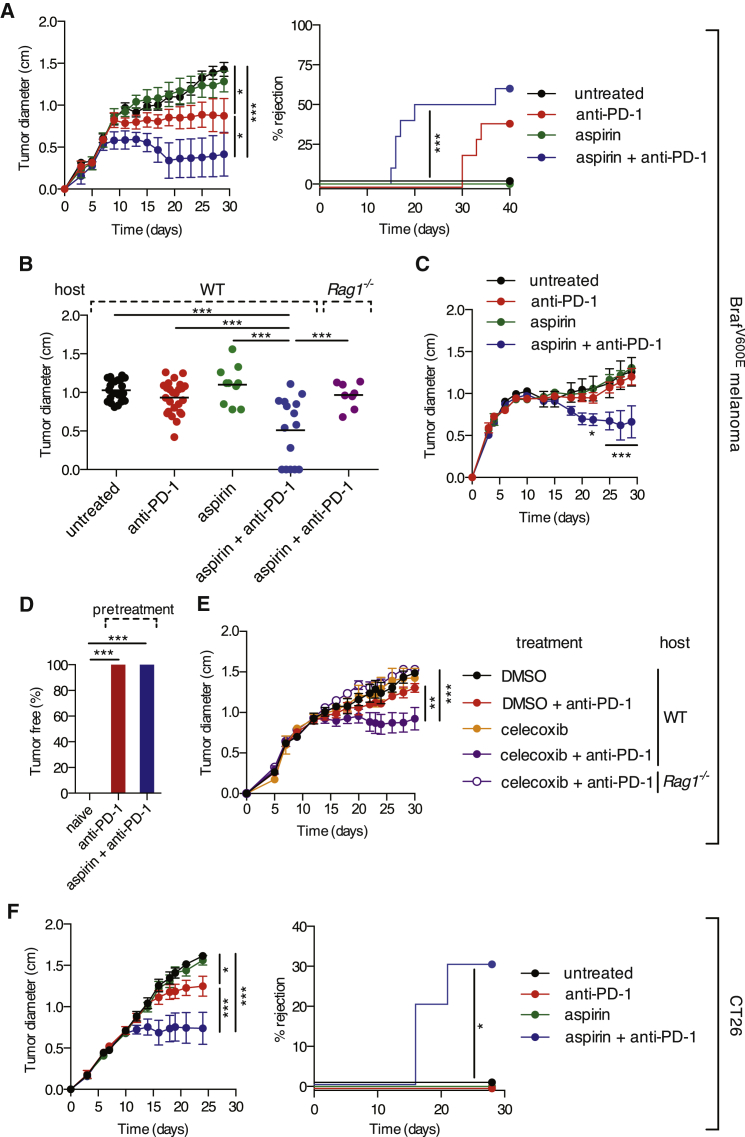
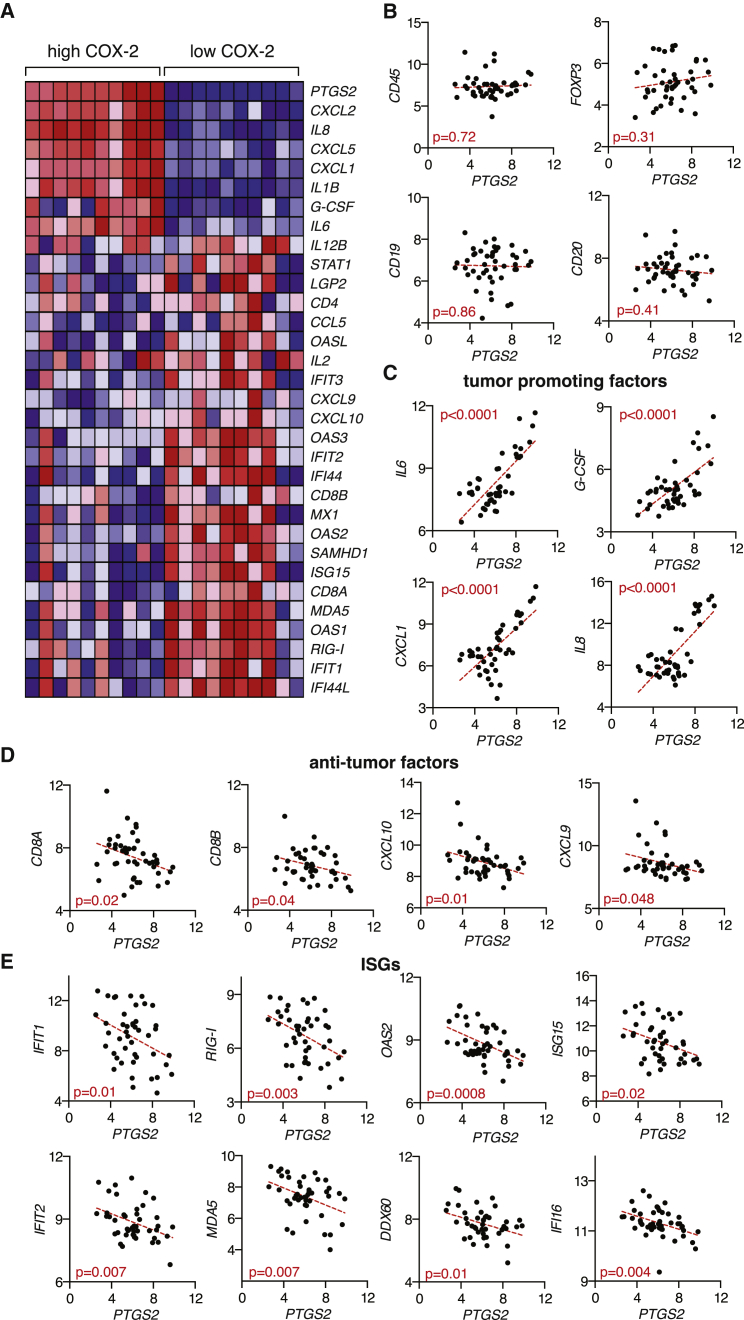
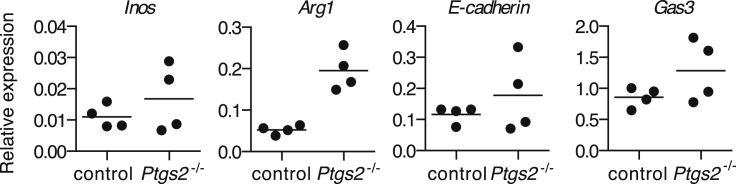
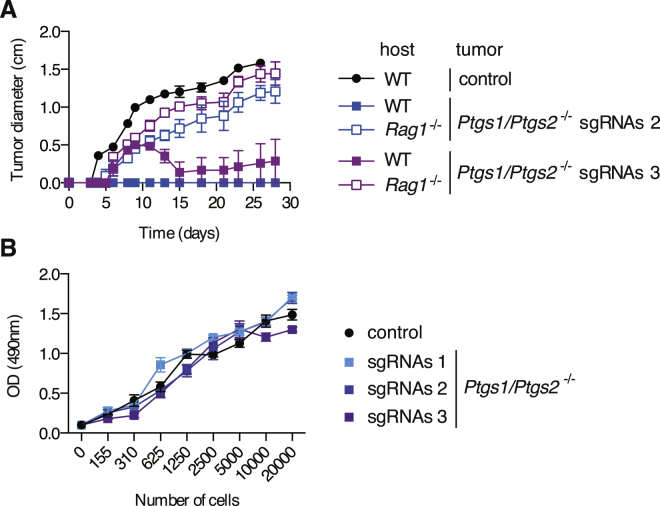
The mechanisms by which melanoma and other cancer cells evade anti-tumor immunity remain incompletely understood. Here, we show that the growth of tumors formed by mutant Braf(V600E) mouse melanoma cells in an immunocompetent host requires their production of prostaglandin E2, which suppresses immunity and fuels tumor-promoting inflammation. Genetic ablation of cyclooxygenases (COX) or prostaglandin E synthases in Braf(V600E) mouse melanoma cells, as well as in Nras(G12D) melanoma or in breast or colorectal cancer cells, renders them susceptible to immune control and provokes a shift in the tumor inflammatory profile toward classic anti-cancer immune pathways. This mouse COX-dependent inflammatory signature is remarkably conserved in human cutaneous melanoma biopsies, arguing for COX activity as a driver of immune suppression across species. Pre-clinical data demonstrate that inhibition of COX synergizes with anti-PD-1 blockade in inducing eradication of tumors, implying that COX inhibitors could be useful adjuvants for immune-based therapies in cancer patients.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
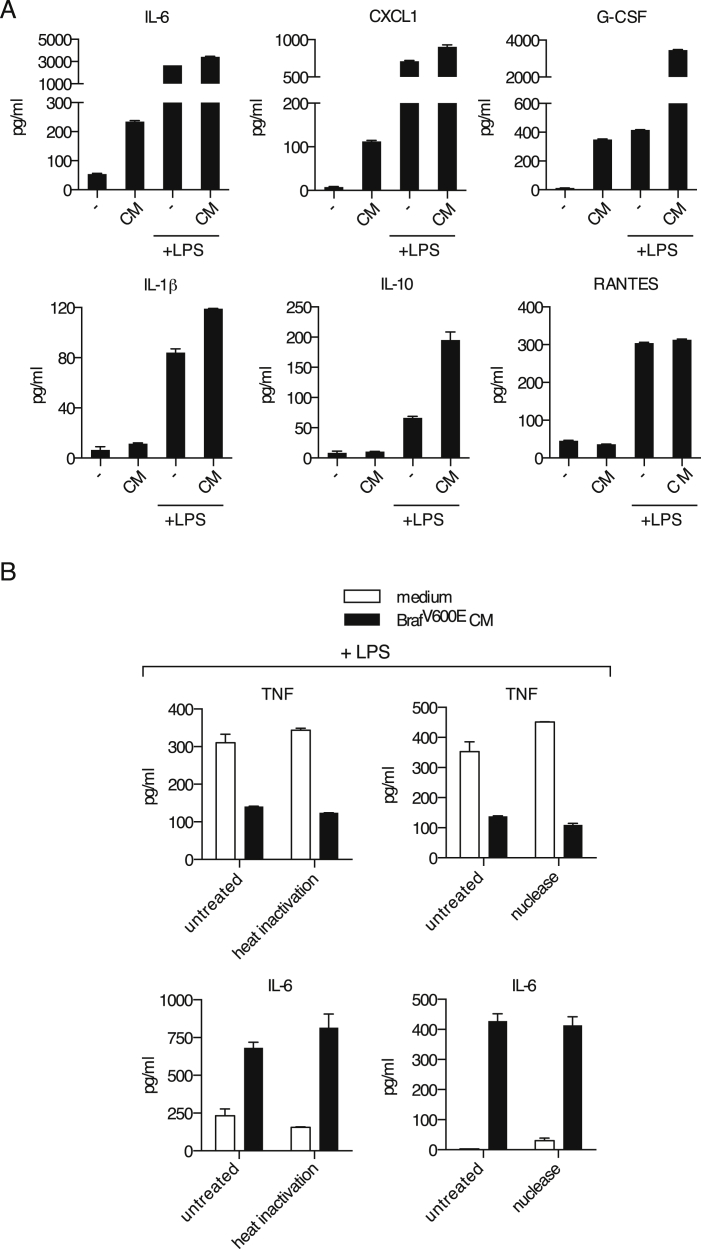
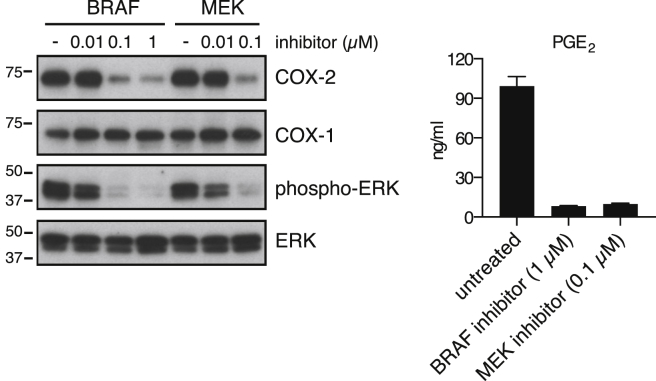
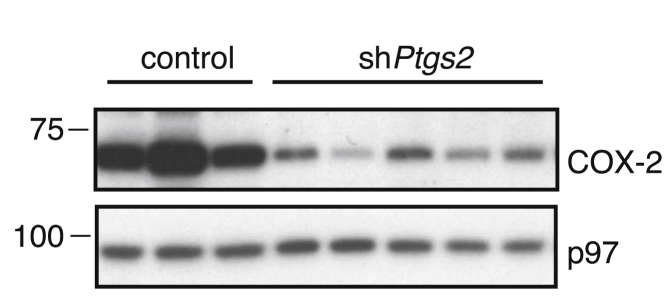
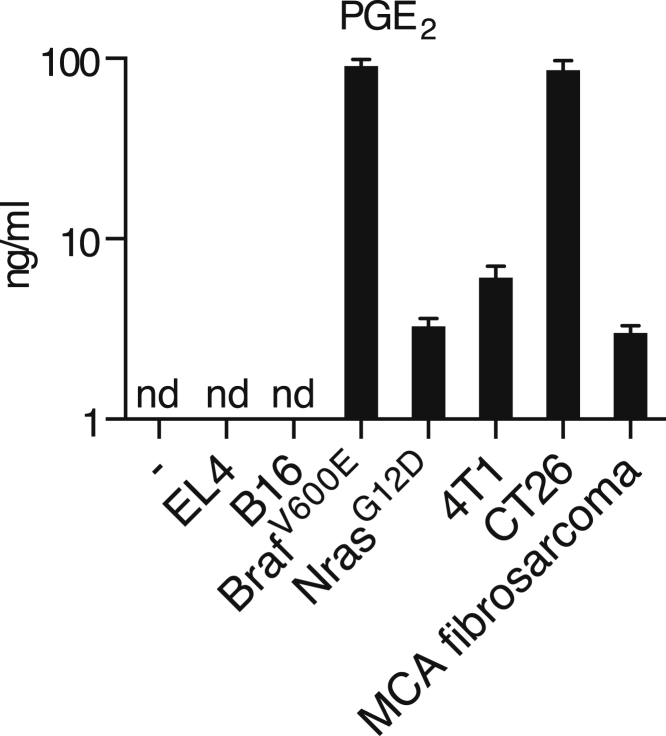
Comment in
-
Immunotherapy: Evading immune escape: synergy of COX and immune-checkpoint inhibitors.Nat Rev Clin Oncol. 2015 Nov;12(11):622. doi: 10.1038/nrclinonc.2015.167. Epub 2015 Sep 22. Nat Rev Clin Oncol. 2015. PMID: 26391776 No abstract available.
-
The Role of Prostaglandin E(2) in Tumor-Associated Immunosuppression.Trends Mol Med. 2016 Jan;22(1):1-3. doi: 10.1016/j.molmed.2015.11.003. Epub 2015 Dec 17. Trends Mol Med. 2016. PMID: 26711015 Free PMC article.
References
-
- Bald T., Landsberg J., Lopez-Ramos D., Renn M., Glodde N., Jansen P., Gaffal E., Steitz J., Tolba R., Kalinke U. Immune cell-poor melanomas benefit from PD-1 blockade after targeted type I IFN activation. Cancer Discov. 2014;4:674–687. - PubMed
-
- Balkwill F., Charles K.A., Mantovani A. Smoldering and polarized inflammation in the initiation and promotion of malignant disease. Cancer Cell. 2005;7:211–217. - PubMed
-
- Becker M.R., Siegelin M.D., Rompel R., Enk A.H., Gaiser T. COX-2 expression in malignant melanoma: a novel prognostic marker? Melanoma Res. 2009;19:8–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
