

Effects of Linker Length and Transient Secondary Structure Elements in the Intrinsically Disordered Notch RAM Region on Notch Signaling
- PMID: 26344835
- PMCID: PMC5576018
- DOI: 10.1016/j.jmb.2015.09.001
Effects of Linker Length and Transient Secondary Structure Elements in the Intrinsically Disordered Notch RAM Region on Notch Signaling
Abstract
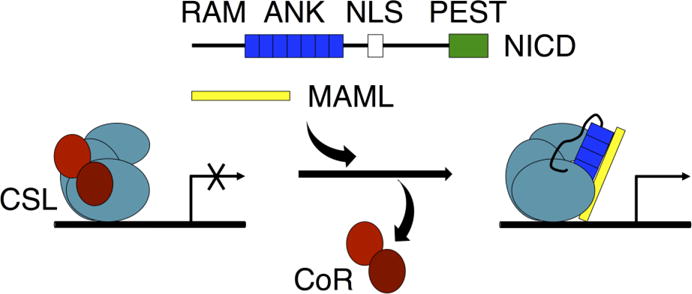
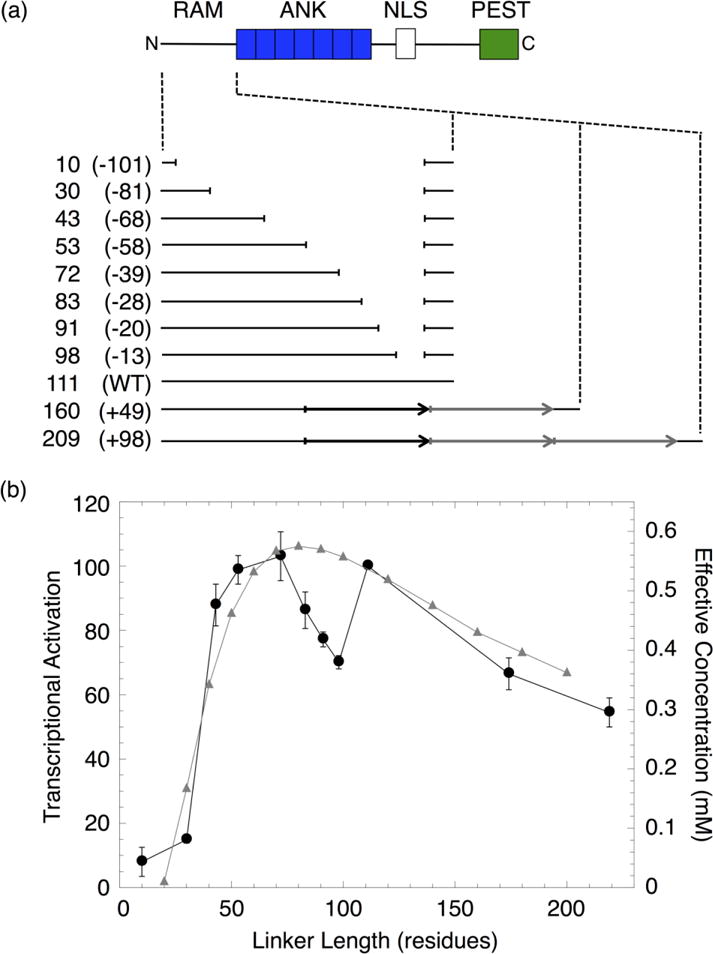
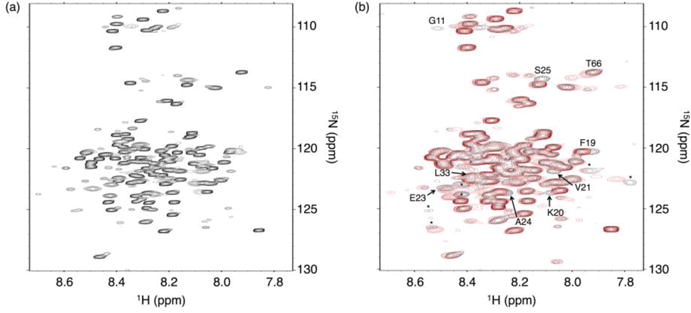
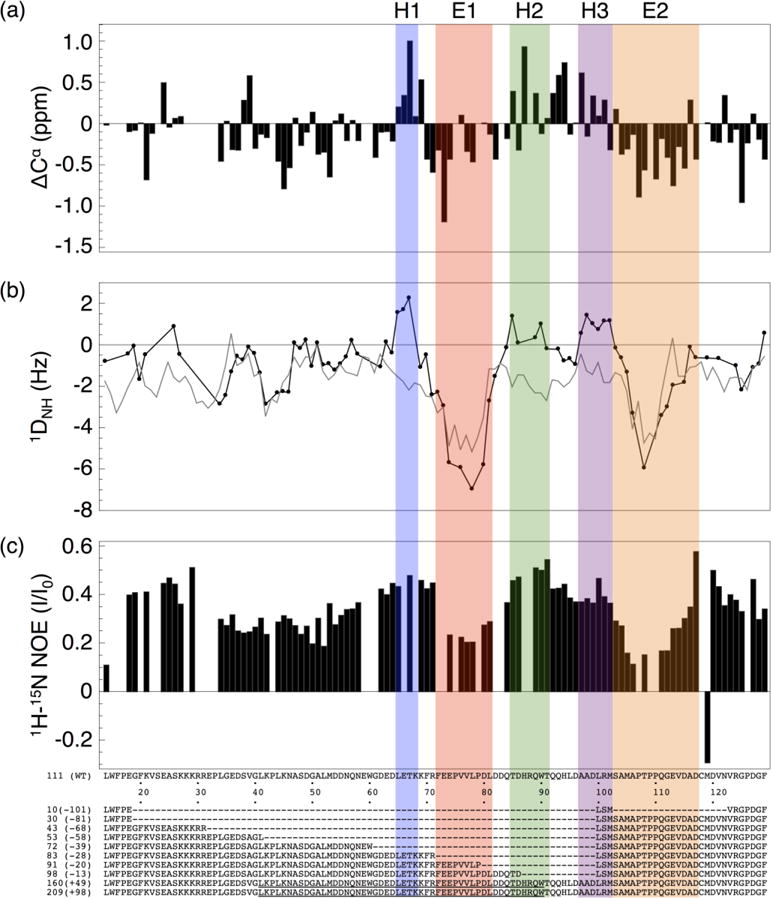
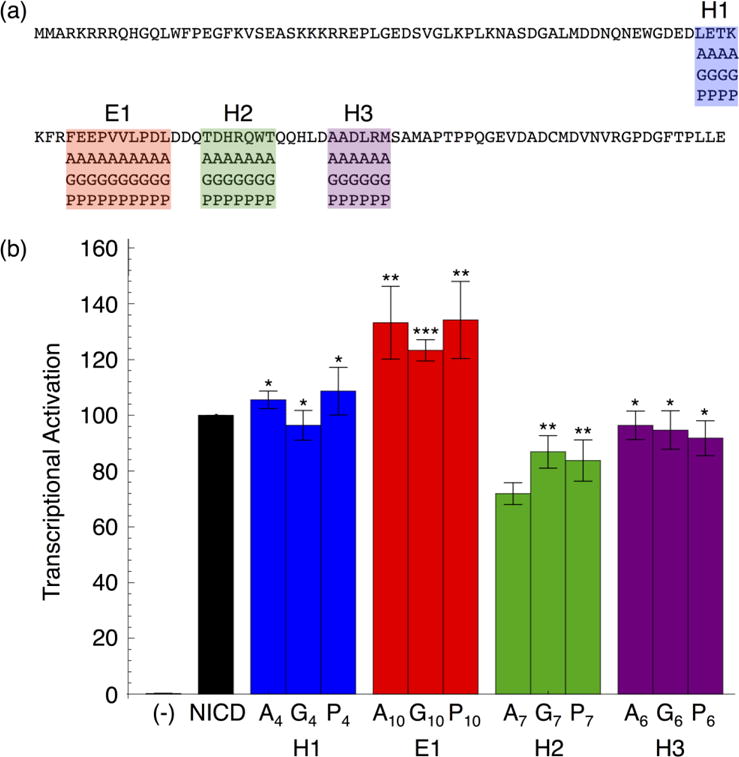
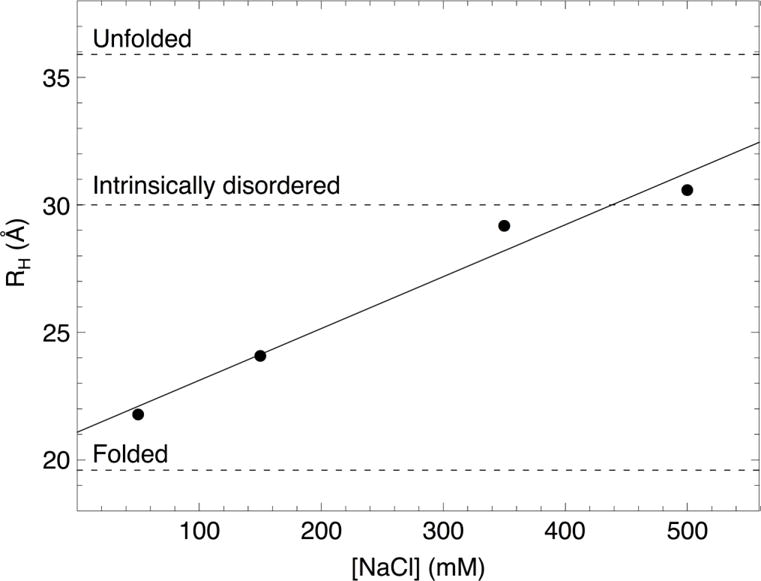
Formation of the bivalent interaction between the Notch intracellular domain (NICD) and the transcription factor CBF-1/RBP-j, Su(H), Lag-1 (CSL) is a key event in Notch signaling because it switches Notch-responsive genes from a repressed state to an activated state. Interaction of the intrinsically disordered RBP-j-associated molecule (RAM) region of NICD with CSL is thought to both disrupt binding of corepressor proteins to CSL and anchor NICD to CSL, promoting interaction of the ankyrin domain of NICD with CSL through an effective concentration mechanism. To quantify the role of disorder in the RAM linker region on the effective concentration enhancement of Notch transcriptional activation, we measured the effects of linker length variation on activation. The resulting activation profile has general features of a worm-like chain model for effective concentration. However, deviations from the model for short sequence deletions suggest that RAM contains sequence-specific structural elements that may be important for activation. Structural characterization of the RAM linker with sedimentation velocity analytical ultracentrifugation and NMR spectroscopy reveals that the linker is compact and contains three transient helices and two extended and dynamic regions. To test if these secondary structure elements are important for activation, we made sequence substitutions to change the secondary structure propensities of these elements and measured transcriptional activation of the resulting variants. Substitutions to two of these nonrandom elements (helix 2, extended region 1) have effects on activation, but these effects do not depend on the nature of the substituting residues. Thus, the primary sequences of these elements, but not their secondary structures, are influencing signaling.
Keywords: NMR spectroscopy; analytical ultracentrifugation; intrinsically disordered proteins; protein–protein interactions; reporter gene assays.
Copyright © 2015. Published by Elsevier Ltd.
Figures
References
-
- Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999;284:770–6. - PubMed
-
- Gridley T. Notch signaling and inherited disease syndromes. Hum Mol Genet. 2003;12:9R–13. - PubMed
-
- Louvi A, Arboleda-Velasquez J, Artavanis-Tsakonas S. CADASIL: a critical look at a Notch disease. Dev Neurosci. 2006;28:5–12. - PubMed
-
- Weng AP, Ferrando AA, Lee W, Morris JP, Silverman LB, Sanchez-Irizarry C, Blacklow SC, Look AT, Aster JC. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia. Science. 2004;306:269–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
