

Elucidating the molecular architecture of adaptation via evolve and resequence experiments
- PMID: 26347030
- PMCID: PMC4733663
- DOI: 10.1038/nrg3937
Elucidating the molecular architecture of adaptation via evolve and resequence experiments
Abstract
Evolve and resequence (E&R) experiments use experimental evolution to adapt populations to a novel environment, then next-generation sequencing to analyse genetic changes. They enable molecular evolution to be monitored in real time on a genome-wide scale. Here, we review the field of E&R experiments across diverse systems, ranging from simple non-living RNA to bacteria, yeast and the complex multicellular organism Drosophila melanogaster. We explore how different evolutionary outcomes in these systems are largely consistent with common population genetics principles. Differences in outcomes across systems are largely explained by different starting population sizes, levels of pre-existing genetic variation, recombination rates and adaptive landscapes. We highlight emerging themes and inconsistencies that future experiments must address.
Figures
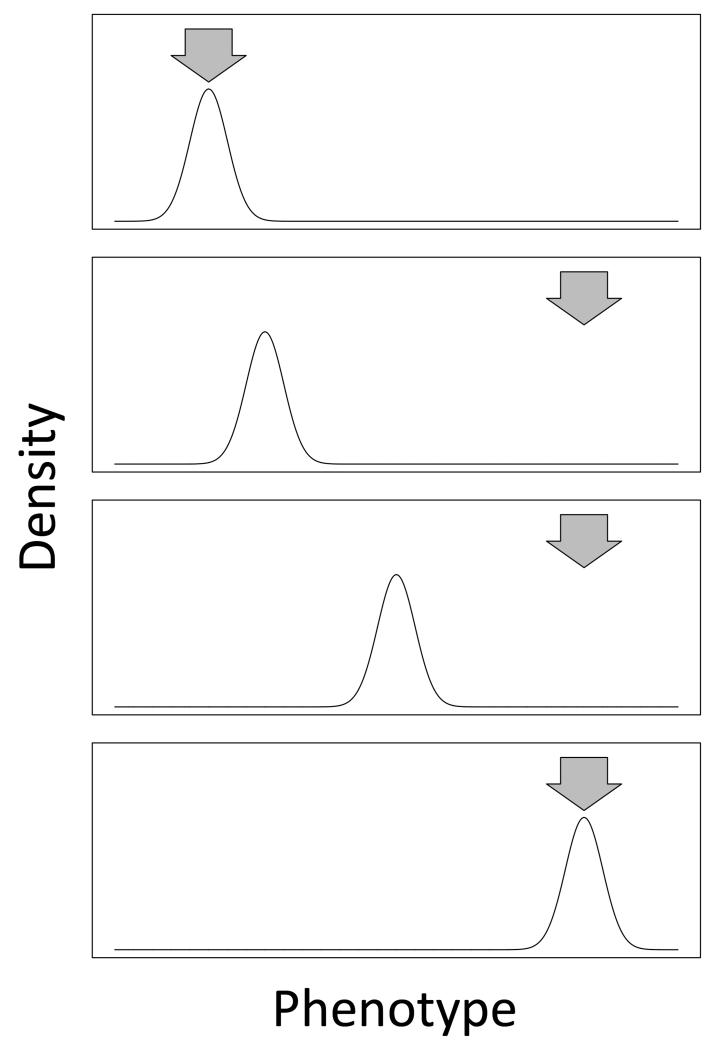
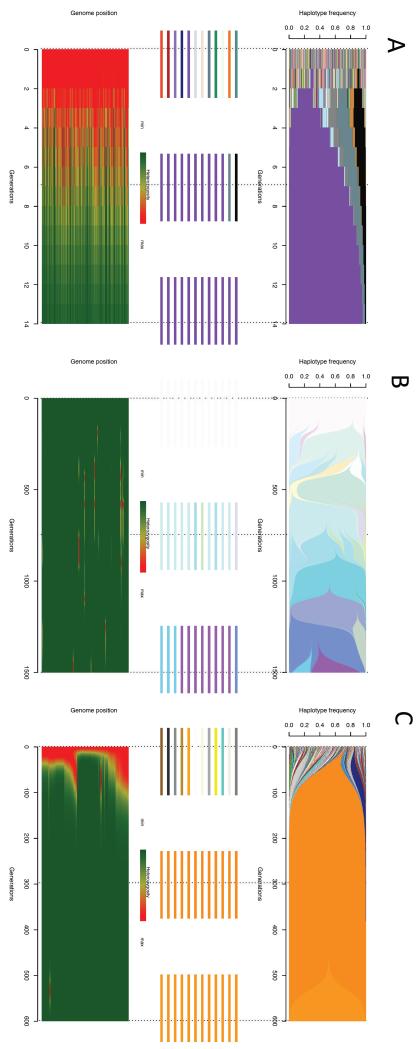
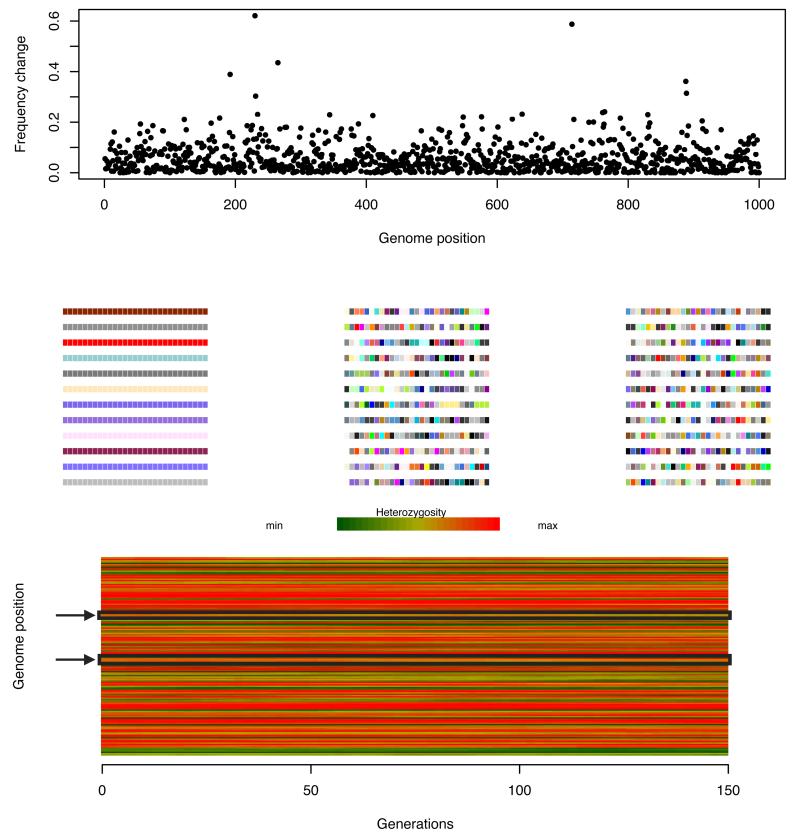
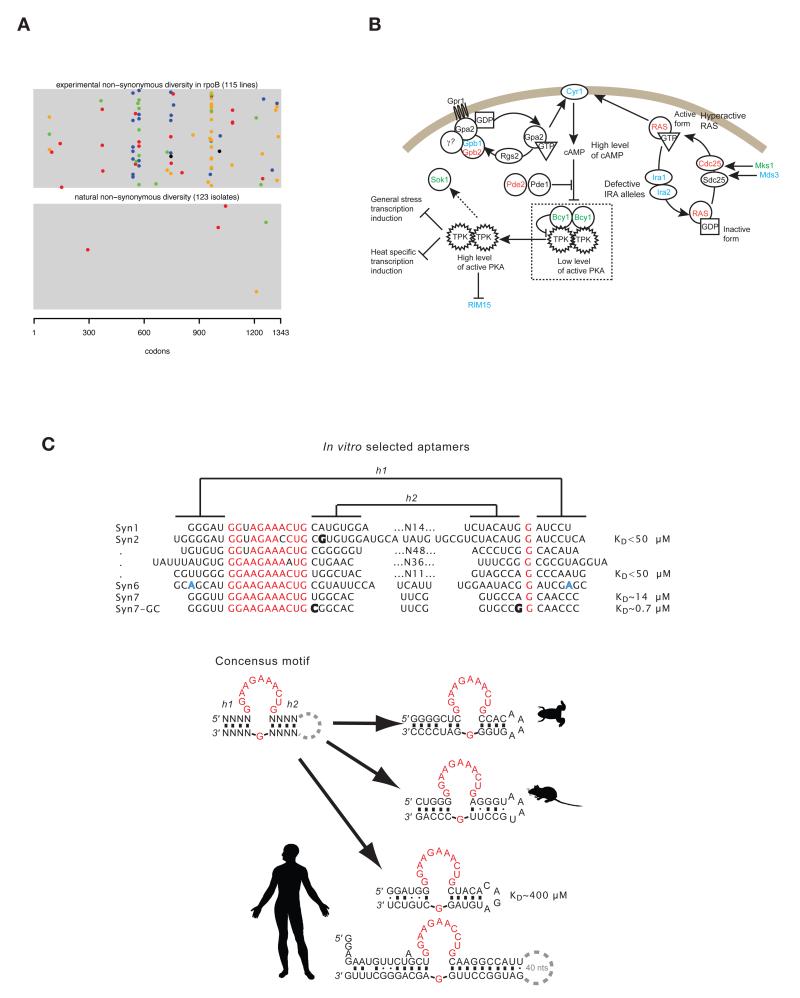
Similar articles
-
Clear: Composition of Likelihoods for Evolve and Resequence Experiments.Genetics. 2017 Jun;206(2):1011-1023. doi: 10.1534/genetics.116.197566. Epub 2017 Apr 10. Genetics. 2017. PMID: 28396506 Free PMC article.
-
Recombination Alters the Dynamics of Adaptation on Standing Variation in Laboratory Yeast Populations.Mol Biol Evol. 2018 Jan 1;35(1):180-201. doi: 10.1093/molbev/msx278. Mol Biol Evol. 2018. PMID: 29069452 Free PMC article.
-
Experimental Evolution as a High-Throughput Screen for Genetic Adaptations.mSphere. 2018 May 9;3(3):e00121-18. doi: 10.1128/mSphere.00121-18. eCollection 2018 May-Jun. mSphere. 2018. PMID: 29743200 Free PMC article.
-
Combining experimental evolution with next-generation sequencing: a powerful tool to study adaptation from standing genetic variation.Heredity (Edinb). 2015 May;114(5):431-40. doi: 10.1038/hdy.2014.86. Epub 2014 Oct 1. Heredity (Edinb). 2015. PMID: 25269380 Free PMC article. Review.
-
Genomic investigations of evolutionary dynamics and epistasis in microbial evolution experiments.Curr Opin Genet Dev. 2015 Dec;35:33-9. doi: 10.1016/j.gde.2015.08.008. Epub 2015 Sep 14. Curr Opin Genet Dev. 2015. PMID: 26370471 Free PMC article. Review.
Cited by
-
The genetic architecture of repeated local adaptation to climate in distantly related plants.Nat Ecol Evol. 2024 Oct;8(10):1933-1947. doi: 10.1038/s41559-024-02514-5. Epub 2024 Aug 26. Nat Ecol Evol. 2024. PMID: 39187610 Free PMC article.
-
The search for sexually antagonistic genes: Practical insights from studies of local adaptation and statistical genomics.Evol Lett. 2020 Aug 31;4(5):398-415. doi: 10.1002/evl3.192. eCollection 2020 Oct. Evol Lett. 2020. PMID: 33014417 Free PMC article.
-
Shared Molecular Targets Confer Resistance over Short and Long Evolutionary Timescales.Mol Biol Evol. 2019 Apr 1;36(4):691-708. doi: 10.1093/molbev/msz006. Mol Biol Evol. 2019. PMID: 30657986 Free PMC article.
-
Laboratory evolution of synthetic electron transport system variants reveals a larger metabolic respiratory system and its plasticity.Nat Commun. 2022 Jun 27;13(1):3682. doi: 10.1038/s41467-022-30877-5. Nat Commun. 2022. PMID: 35760776 Free PMC article.
-
Selection and the direction of phenotypic evolution.Elife. 2023 Aug 31;12:e80993. doi: 10.7554/eLife.80993. Elife. 2023. PMID: 37650381 Free PMC article.
References
-
- Adams J, Rosenzweig F. Experimental microbial evolution: history and conceptual underpinnings. Genomics. 2014;104:393–398. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
