

The allosteric behavior of Fur mediates oxidative stress signal transduction in Helicobacter pylori
- PMID: 26347726
- PMCID: PMC4541418
- DOI: 10.3389/fmicb.2015.00840
The allosteric behavior of Fur mediates oxidative stress signal transduction in Helicobacter pylori
Abstract
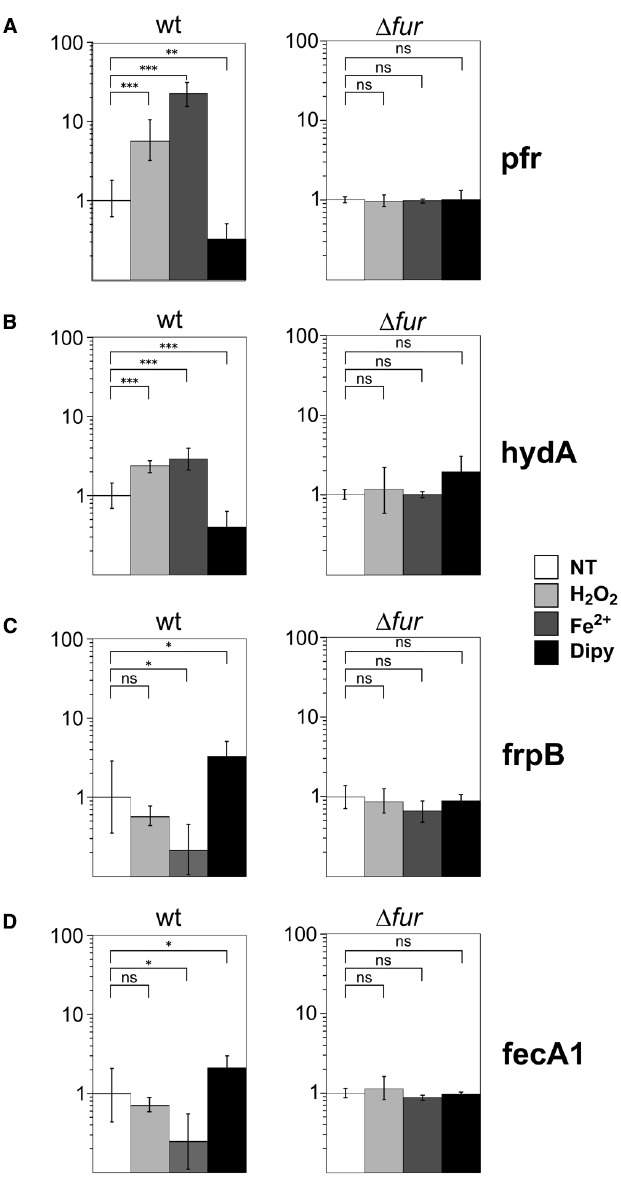
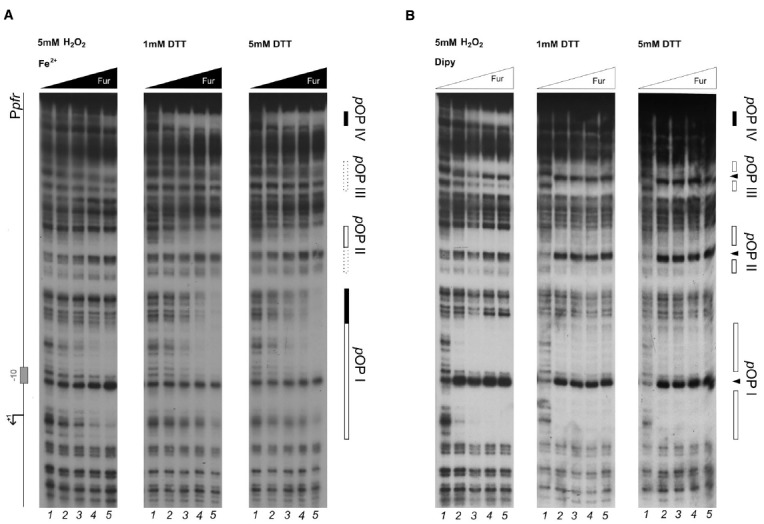
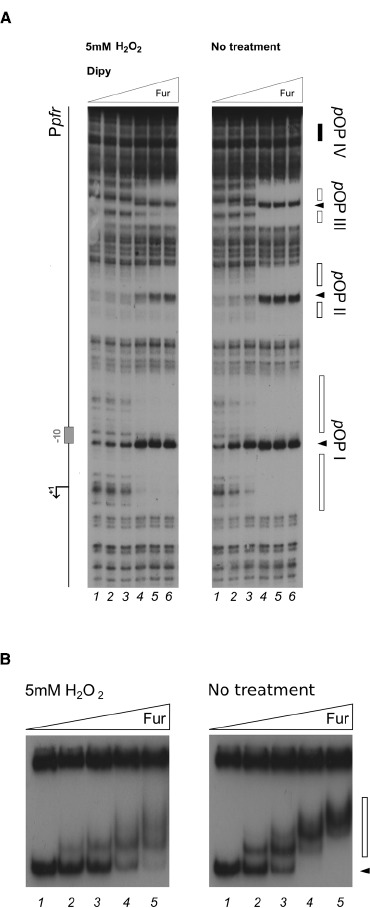
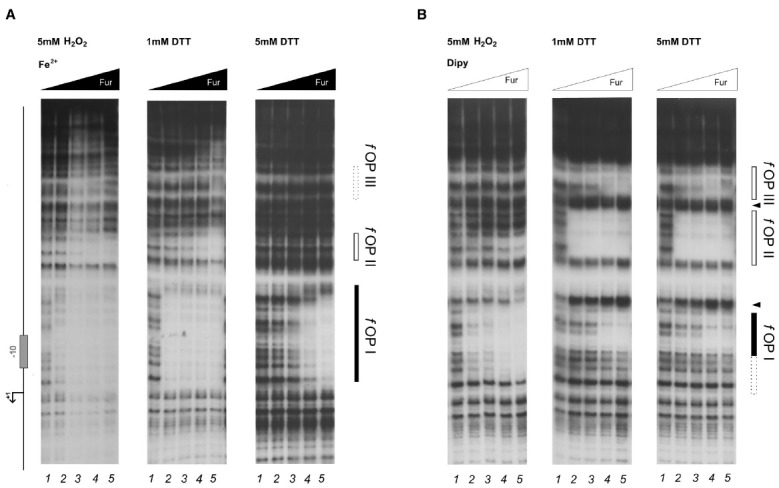
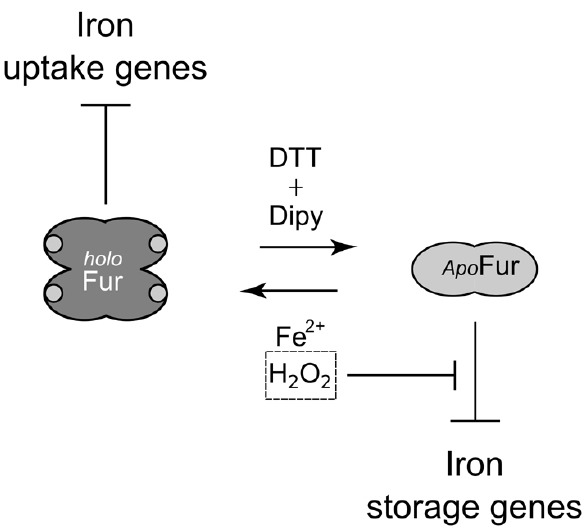
The microaerophilic gastric pathogen Helicobacter pylori is exposed to oxidative stress originating from the aerobic environment, the oxidative burst of phagocytes and the formation of reactive oxygen species, catalyzed by iron excess. Accordingly, the expression of genes involved in oxidative stress defense have been repeatedly linked to the ferric uptake regulator Fur. Moreover, mutations in the Fur protein affect the resistance to metronidazole, likely due to loss-of-function in the regulation of genes involved in redox control. Although many advances in the molecular understanding of HpFur function were made, little is known about the mechanisms that enable Fur to mediate the responses to oxidative stress. Here we show that iron-inducible, apo-Fur repressed genes, such as pfr and hydA, are induced shortly after oxidative stress, while their oxidative induction is lost in a fur knockout strain. On the contrary, holo-Fur repressed genes, such as frpB1 and fecA1, vary modestly in response to oxidative stress. This indicates that the oxidative stress signal specifically targets apo-Fur repressed genes, rather than impairing indiscriminately the regulatory function of Fur. Footprinting analyses showed that the oxidative signal strongly impairs the binding affinity of Fur toward apo-operators, while the binding toward holo-operators is less affected. Further evidence is presented that a reduced state of Fur is needed to maintain apo-repression, while oxidative conditions shift the preferred binding architecture of Fur toward the holo-operator binding conformation, even in the absence of iron. Together the results demonstrate that the allosteric regulation of Fur enables transduction of oxidative stress signals in H. pylori, supporting the concept that apo-Fur repressed genes can be considered oxidation inducible Fur regulatory targets. These findings may have important implications in the study of H. pylori treatment and resistance to antibiotics.
Keywords: allosteric regulation; antibiotic resistance; ferric uptake regulator; metal homeostasis; metalloproteins; oxidative stress; redox regulation; transcriptional regulation.
Figures

Similar articles
-
Identification and characterization of novel Helicobacter pylori apo-fur-regulated target genes.J Bacteriol. 2013 Dec;195(24):5526-39. doi: 10.1128/JB.01026-13. Epub 2013 Oct 4. J Bacteriol. 2013. PMID: 24097951 Free PMC article.
-
(Re)-definition of the holo- and apo-Fur direct regulons of Helicobacter pylori.J Mol Biol. 2024 May 15;436(10):168573. doi: 10.1016/j.jmb.2024.168573. Epub 2024 Apr 16. J Mol Biol. 2024. PMID: 38626867
-
Random and site-specific mutagenesis of the Helicobacter pylori ferric uptake regulator provides insight into Fur structure-function relationships.Mol Microbiol. 2013 Jul;89(2):304-23. doi: 10.1111/mmi.12278. Epub 2013 Jun 10. Mol Microbiol. 2013. PMID: 23710935 Free PMC article.
-
Redox Sensing by Fe2+ in Bacterial Fur Family Metalloregulators.Antioxid Redox Signal. 2018 Dec 20;29(18):1858-1871. doi: 10.1089/ars.2017.7359. Epub 2017 Oct 31. Antioxid Redox Signal. 2018. PMID: 28938859 Free PMC article. Review.
-
How we learnt about iron acquisition in Pseudomonas aeruginosa: a series of very fortunate events.Biometals. 2007 Jun;20(3-4):587-601. doi: 10.1007/s10534-006-9067-2. Epub 2006 Dec 22. Biometals. 2007. PMID: 17186376 Review.
Cited by
-
The Helicobacter pylori Heat-Shock Repressor HspR: Definition of Its Direct Regulon and Characterization of the Cooperative DNA-Binding Mechanism on Its Own Promoter.Front Microbiol. 2018 Aug 14;9:1887. doi: 10.3389/fmicb.2018.01887. eCollection 2018. Front Microbiol. 2018. PMID: 30154784 Free PMC article.
-
BlsA integrates light and temperature signals into iron metabolism through Fur in the human pathogen Acinetobacter baumannii.Sci Rep. 2018 May 16;8(1):7728. doi: 10.1038/s41598-018-26127-8. Sci Rep. 2018. PMID: 29769610 Free PMC article.
-
Comprehensive mapping of the Helicobacter pylori NikR regulon provides new insights in bacterial nickel responses.Sci Rep. 2017 Apr 10;7:45458. doi: 10.1038/srep45458. Sci Rep. 2017. PMID: 28393877 Free PMC article.
-
RNase Y mediates posttranscriptional control of the virulence-associated CncR1 small-RNA in Helicobacter pylori.iScience. 2025 Jan 16;28(2):111815. doi: 10.1016/j.isci.2025.111815. eCollection 2025 Feb 21. iScience. 2025. PMID: 39949958 Free PMC article.
-
Antibiotic-induced ROS-mediated Fur allosterism contributes to Helicobacter pylori resistance by inhibiting arsR activation of mutS and mutY.Antimicrob Agents Chemother. 2024 Apr 3;68(4):e0167923. doi: 10.1128/aac.01679-23. Epub 2024 Feb 22. Antimicrob Agents Chemother. 2024. PMID: 38386782 Free PMC article.
References
-
- Agriesti F., Roncarati D., Musiani F., Del Campo C., Iurlaro M., Sparla F., et al. (2014). FeON-FeOFF: the Helicobacter pylori Fur regulator commutates iron-responsive transcription by discriminative readout of opposed DNA grooves. Nucleic Acids Res. 42, 3138–3151. 10.1093/nar/gkt1258 - DOI - PMC - PubMed
-
- Barnard F. M., Loughlin M. F., Fainberg H. P., Messenger M. P., Ussery D. W., Williams P., et al. (2004). Global regulation of virulence and the stress response by CsrA in the highly adapted human gastric pathogen Helicobacter pylori. Mol. Microbiol. 51, 15–32. 10.1046/j.1365-2958.2003.03788.x - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources