

The Chromatin Protein DUET/MMD1 Controls Expression of the Meiotic Gene TDM1 during Male Meiosis in Arabidopsis
- PMID: 26348709
- PMCID: PMC4562639
- DOI: 10.1371/journal.pgen.1005396
The Chromatin Protein DUET/MMD1 Controls Expression of the Meiotic Gene TDM1 during Male Meiosis in Arabidopsis
Abstract
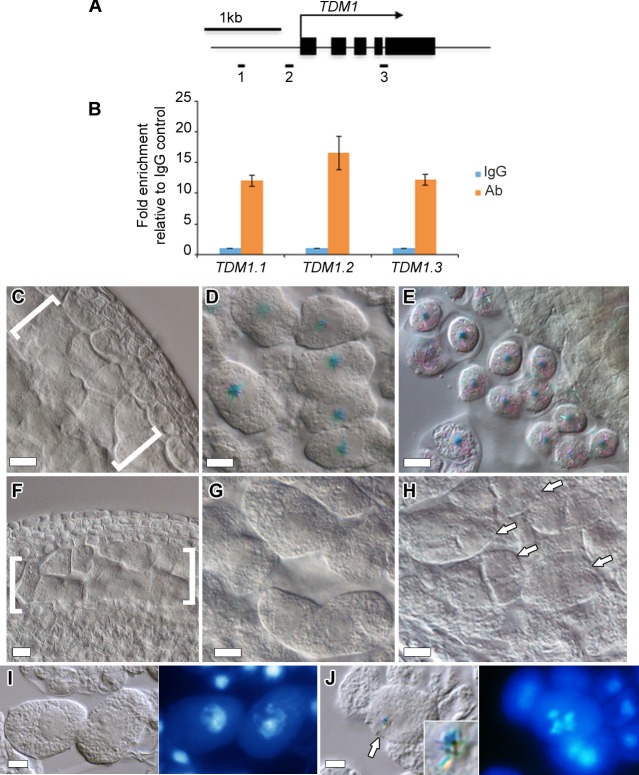
Meiosis produces haploid cells essential for sexual reproduction. In yeast, entry into meiosis activates transcription factors which trigger a transcriptional cascade that results in sequential co-expression of early, middle and late meiotic genes. However, these factors are not conserved, and the factors and regulatory mechanisms that ensure proper meiotic gene expression in multicellular eukaryotes are poorly understood. Here, we report that DUET/MMD1, a PHD finger protein essential for Arabidopsis male meiosis, functions as a transcriptional regulator in plant meiosis. We find that DUET-PHD binds H3K4me2 in vitro, and show that this interaction is critical for function during meiosis. We also show that DUET is required for proper microtubule organization during meiosis II, independently of its function in meiosis I. Remarkably, DUET protein shows stage-specific expression, confined to diplotene. We identify two genes TDM1 and JAS with critical functions in cell cycle transitions and spindle organization in male meiosis, as DUET targets, with TDM1 being a direct target. Thus, DUET is required to regulate microtubule organization and cell cycle transitions during male meiosis, and functions as a direct transcription activator of the meiotic gene TDM1. Expression profiling showed reduced expression of a subset comprising about 12% of a known set of meiosis preferred genes in the duet mutant. Our results reveal the action of DUET as a transcriptional regulator during male meiosis in plants, and suggest that transcription of meiotic genes is under stagewise control in plants as in yeast.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

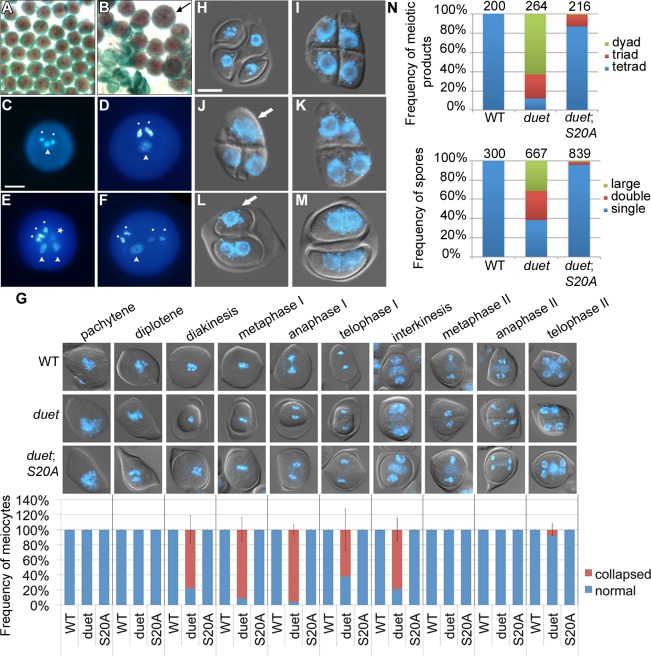
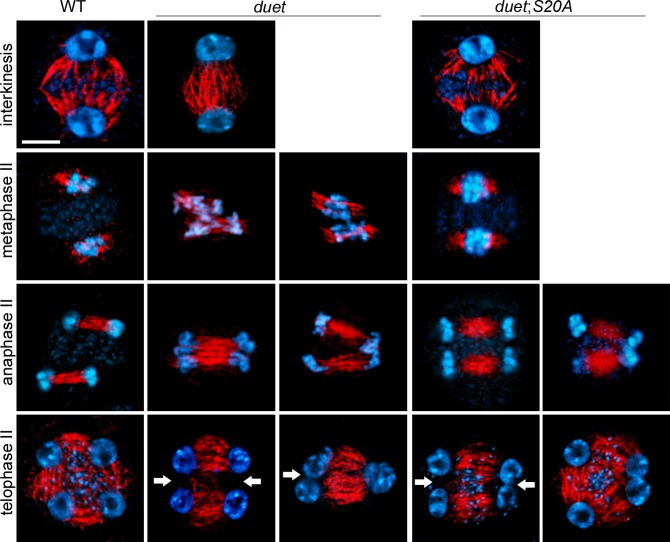
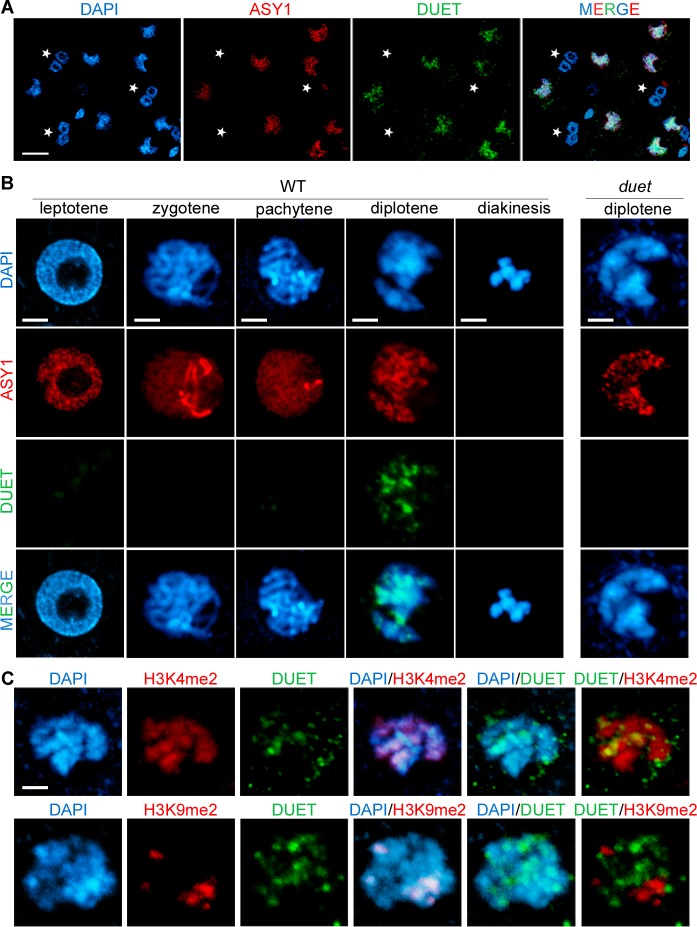
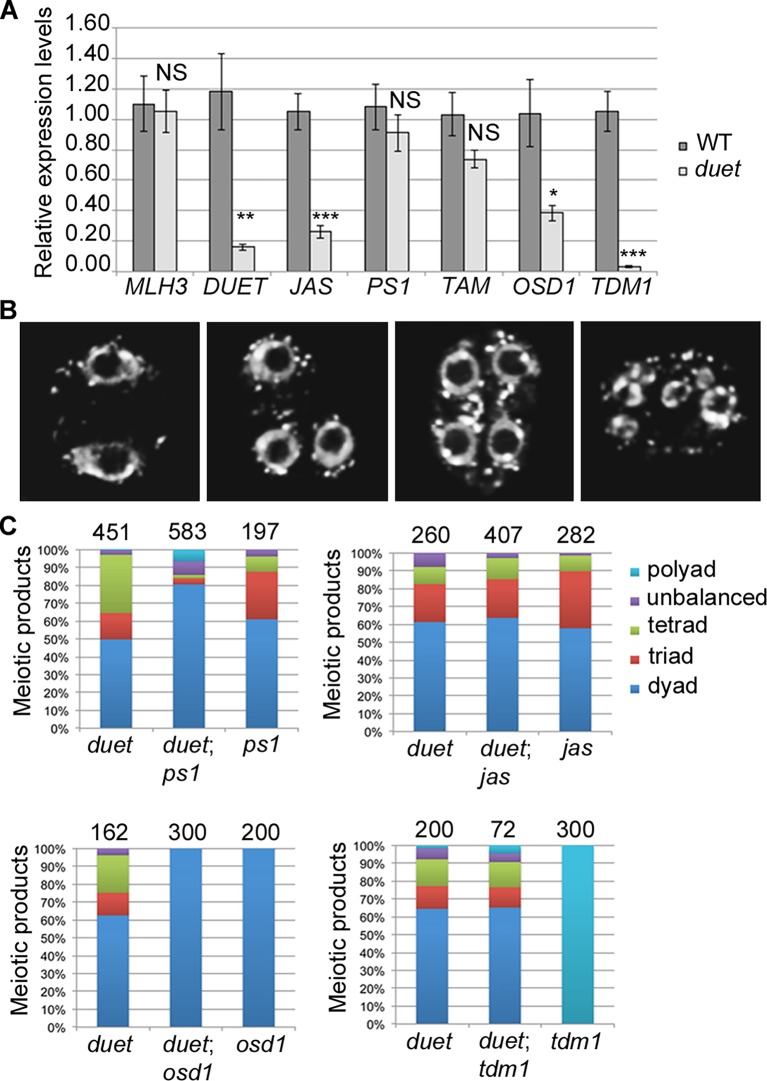
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
