

Review
doi: 10.1101/cshperspect.a020495.
The Nodes of Ranvier: Molecular Assembly and Maintenance
Affiliations
- PMID: 26354894
- PMCID: PMC4772103
- DOI: 10.1101/cshperspect.a020495
Item in Clipboard
Review
The Nodes of Ranvier: Molecular Assembly and Maintenance
Cold Spring Harb Perspect Biol.
.
Abstract
Action potential (AP) propagation in myelinated nerves requires clustered voltage gated sodium and potassium channels. These channels must be specifically localized to nodes of Ranvier where the AP is regenerated. Several mechanisms have evolved to facilitate and ensure the correct assembly and stabilization of these essential axonal domains. This review highlights the current understanding of the axon intrinsic and glial extrinsic mechanisms that control the formation and maintenance of the nodes of Ranvier in both the peripheral nervous system (PNS) and central nervous system (CNS).
Copyright © 2016 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
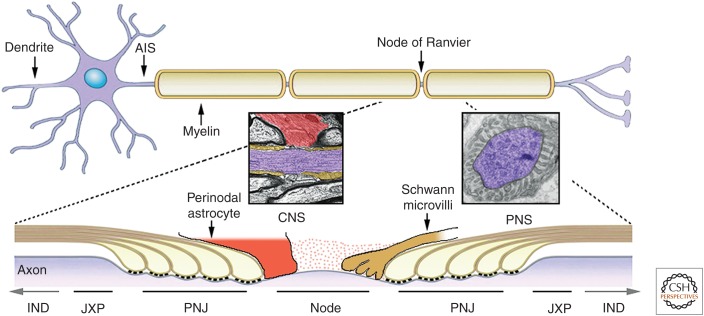
Axonal domains along the myelinated axons. A neuron containing the soma, branched dendrites, and a myelinated axon is shown. Axons are myelinated by Schwann cells in the peripheral nervous system (PNS) and oligodendrocytes in the central nervous system (CNS). Action potentials (APs) generated at the axon initial segment (AIS) travel down the axon and are regenerated at the nodes of Ranvier until reaching the nerve terminals. The axonal membrane is divided into distinct domains. The internodes (INDs) (shown partially in the lower cartoon) comprise the majority of the axon and are located beneath the compact myelin sheath. The juxtaparanodes (JXP) are located at the end of the internodes. Near the nodes of Ranvier, the myelin sheath ends with a series of cytoplasmic loops (e.g., paranodal loops) that generate a specialized junction with the axon (paranodal junction [PNJ], often referred to as the axoglial junction). Bordered by the PNJ are the nodes of Ranvier, which are gaps between myelin segments. In the PNS, the nodal axolemma is contacted by microvilli that originate from the outer aspect of the myelinating Schwann cells, whereas, in the CNS, some nodes are contacted by a process from a perinodal astrocyte or oligodendrocyte progenitor cell (OPC). Insets show EM images of a longitudinal section through the node of Ranvier in the CNS (perinodal astrocyte in red, paranodes in brown, and axon in purple) and the PNS (axon in purple). The nodal gap is filled with highly charged extracellular matrix material (dots).
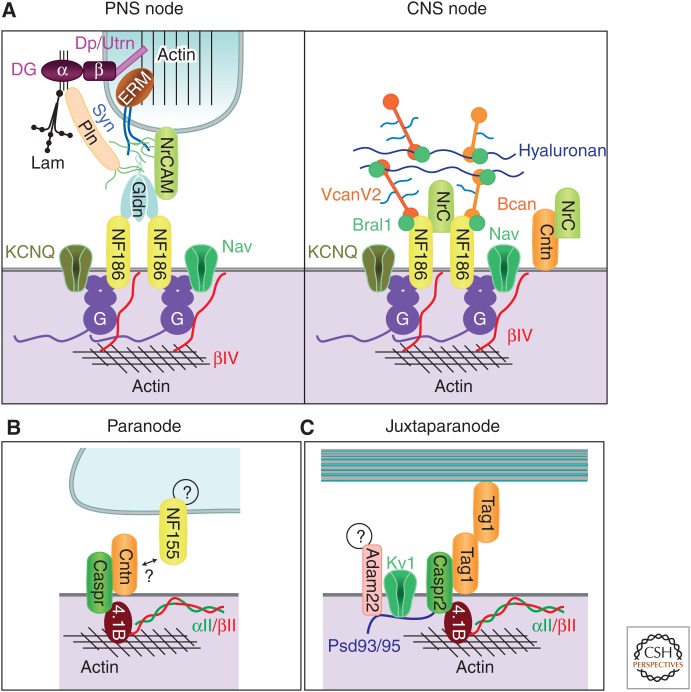
Molecular organizations of axonal subdomains. Simplified illustration showing some of the molecules and interactions involved in the nodal (A), paranodal (B), and juxtaparanodal (C) domains. PNS nodes are contacted by microvilli of Schwann cells. Dystroglycan (DG) is autocleaved into α and β chains, which remain associated. β-DG interacts with dystrophin (Dp) and utrophin (Utrn). A transmembrane form of NrCAM is present at the microvilli. Interaction of laminins (Lam) and perlecan (Pln) with α-DG requires proper glycosylation of α-DG (thin black lines). Pln and sydecans 3/4 (Syn) are modified by heparan sulfate side chains (green lines). The furin-shed gliomedin (Gldn) trimerizes and is associated with heparan sulfate through its amino-terminal region and collagen-like domain and interacts with NrCAM and NF186 through its olfactomedin domain. G and βIV represent AnkG and βIV spectrin, respectively. In the CNS, the nodal ECM is enriched with shed NrCAM, but its cellular source is unknown. VcanV2 and Bcan interact with Bral1 and hyaluronan through their G1 globular domains and with NF186 through the G3 domains. The intervening regions of VcanV2 and Bcan are modified by chondroitin sulfate side chains (light blue lines). Contactin (Cntn) was found at CNS nodes, but only weakly at a few PNS nodes. The cytoplasmic partners of NF155 at the paranodal junction and ligands of Adam22 at the juxtaparanode are currently unknown. (From Chang et al. 2013; reprinted, with permission, from Elsevier Limited © 2013.)
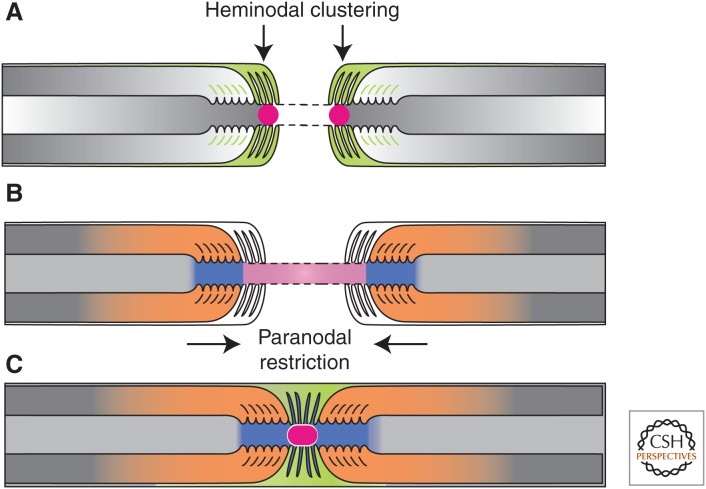
The formation of PNS nodes of Ranvier. (A) Na+ channels (red circle) are trapped at heminodes that are contacted by Schwann cell microvilli ([MV] orange). Axon–glia interaction at this site is mediated by binding of gliomedin and glial NrCAM to axonal NF186. A transmembrane form of glial NrCAM traps gliomedin on Schwann cell microvilli and enhances its binding to axonal NF186. (B) The distribution of Na+ channels is restricted between two forming myelin segments by the PNJ (blue). Three CAMs, NF155 present at the glial paranodal loops, and an axonal complex of Caspr and contactin mediate axon–glia interaction and the formation of the PNJ. (C) These two extrinsic cooperating mechanisms provide reciprocal backup systems and ensure that Na+ channels are found at high density at the nodes. Finally, the entire nodal complex requires stabilization and interaction with cytoskeletal and scaffolding proteins, which constitute a third, intrinsic mechanism for Na+ channel clustering. (From Feinberg et al. 2010; reprinted, with permission, from Elsevier Limited © 2010.)
References
-
- Arroyo EJ, Xu YT, Zhou L, Messing A, Peles E, Chiu SY, Scherer SS. 1999. Myelinating Schwann cells determine the internodal localization of Kv1.1, Kv1.2, Kvβ2, and Caspr. J Neurocytol 28: 333–347. - PubMed
-
- Arroyo EJ, Xu T, Poliak S, Watson M, Peles E, Scherer SS. 2001. Internodal specializations of myelinated axons in the central nervous system. Cell Tissue Res 305: 53–66. - PubMed
-
- Bangratz M, Sarrazin N, Devaux J, Zambroni D, Echaniz-Laguna A, René F, Boërio D, Davoine CS, Fontaine B, Feltri ML, et al. 2012. A mouse model of Schwartz–Jampel syndrome reveals myelinating Schwann cell dysfunction with persistent axonal depolarization in vitro and distal peripheral nerve hyperexcitability when perlecan is lacking. Am J Pathol 180: 2040–2055. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources