

Catecholaminergic Fiber Innervation of the Vocal Motor System Is Intrasexually Dimorphic in a Teleost with Alternative Reproductive Tactics
- PMID: 26355302
- PMCID: PMC4745882
- DOI: 10.1159/000438720
Catecholaminergic Fiber Innervation of the Vocal Motor System Is Intrasexually Dimorphic in a Teleost with Alternative Reproductive Tactics
Abstract
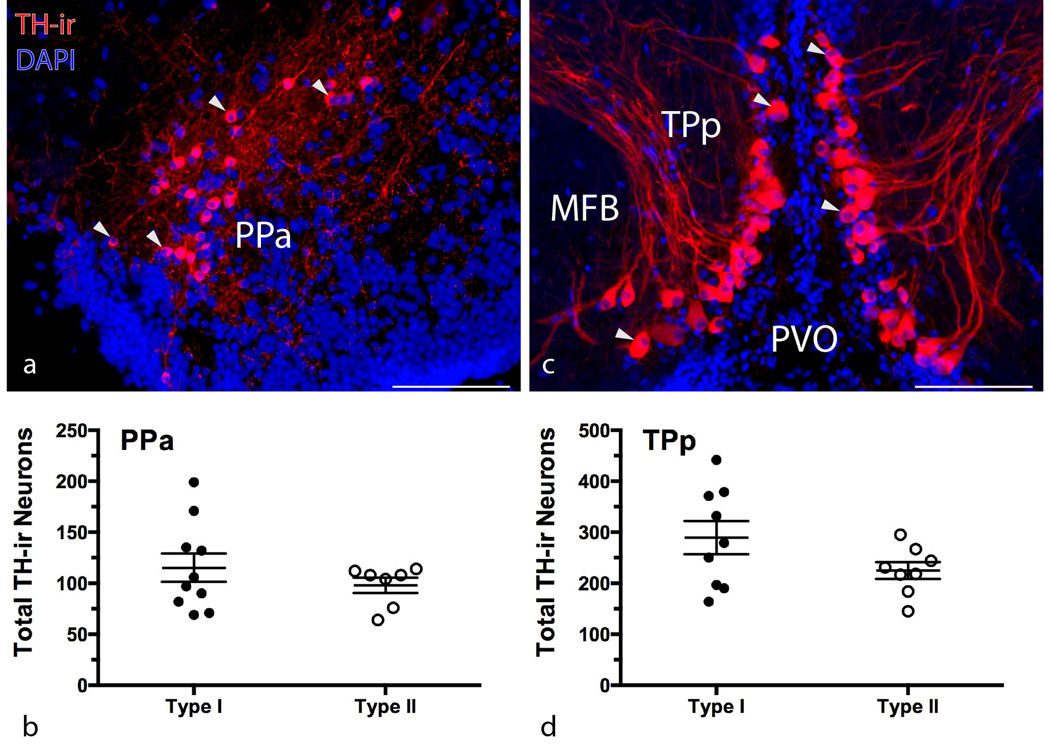
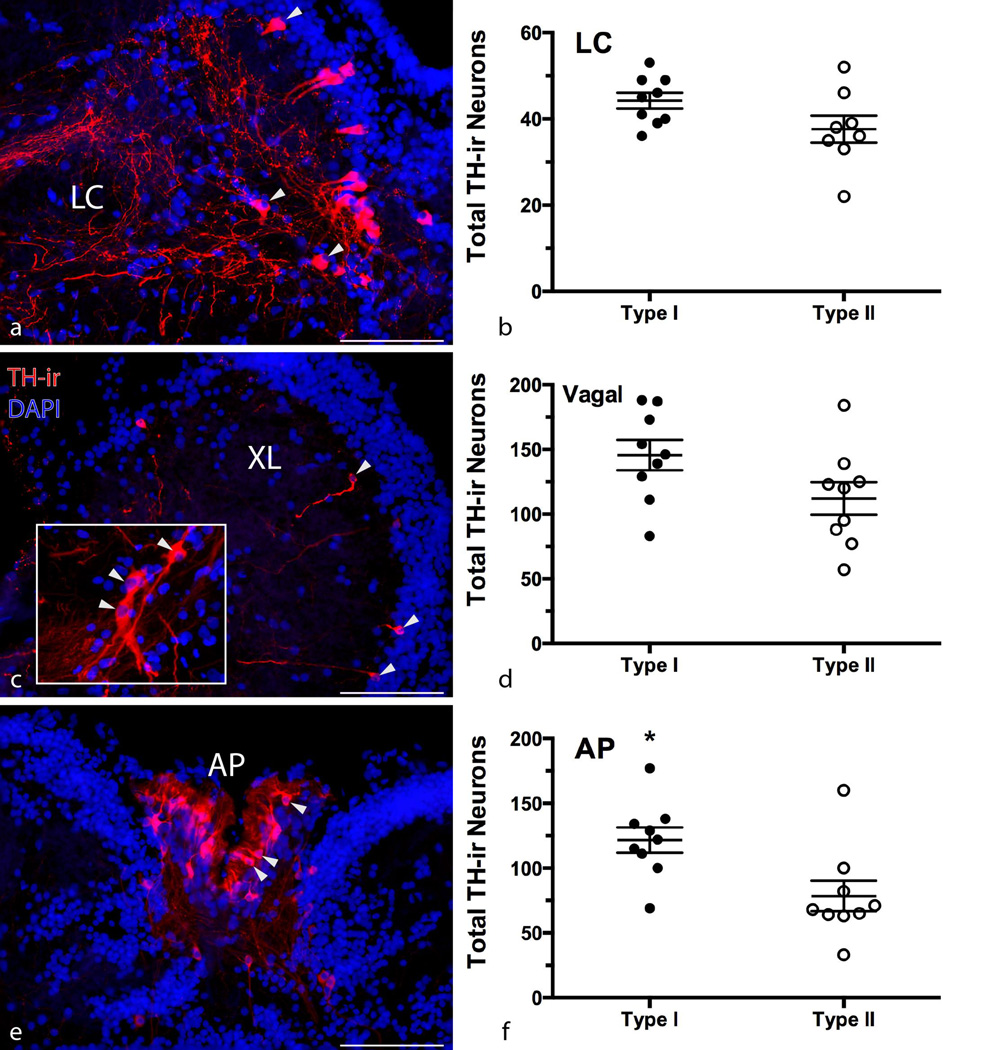
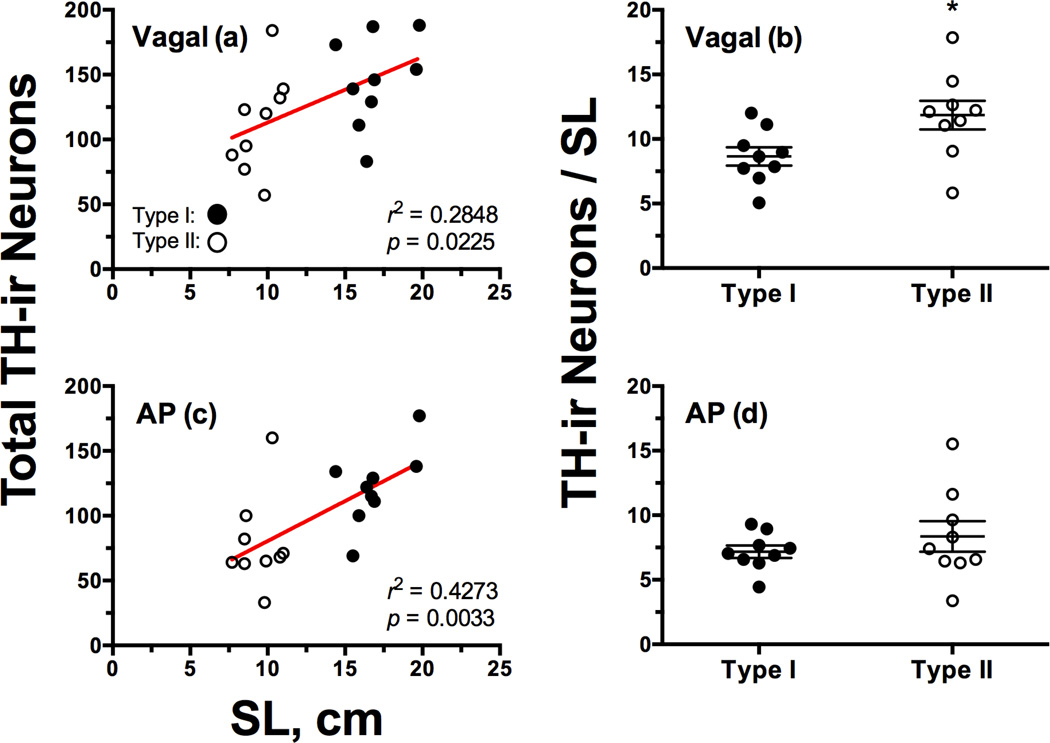
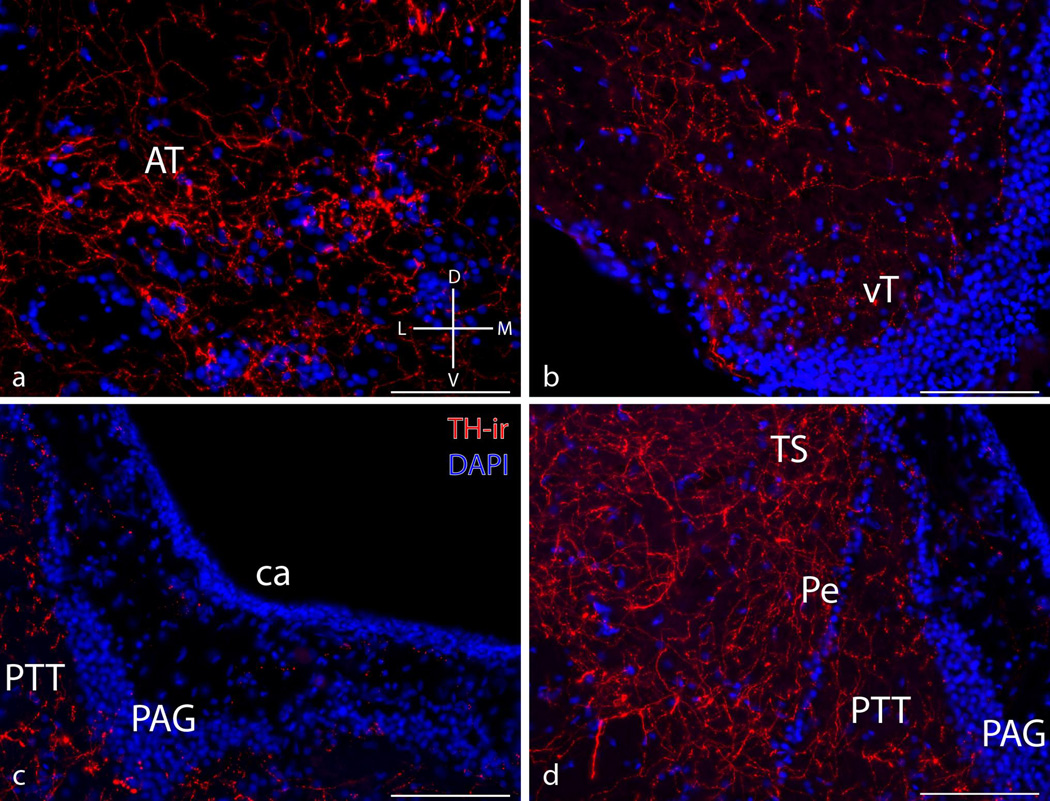
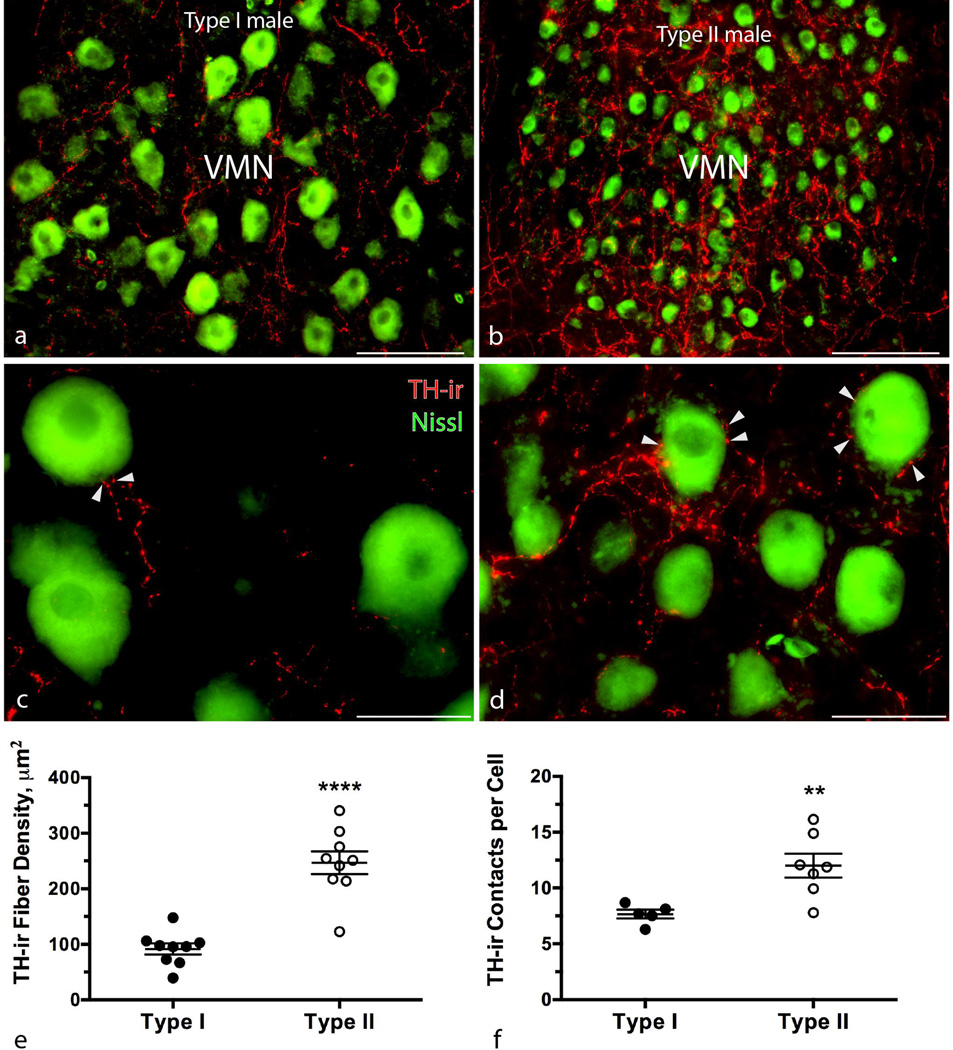
Catecholamines, which include the neurotransmitters dopamine and noradrenaline, are known modulators of sensorimotor function, reproduction, and sexually motivated behaviors across vertebrates, including vocal-acoustic communication. Recently, we demonstrated robust catecholaminergic (CA) innervation throughout the vocal motor system in the plainfin midshipman fish Porichthys notatus, a seasonal breeding marine teleost that produces vocal signals for social communication. There are 2 distinct male reproductive morphs in this species: type I males establish nests and court females with a long-duration advertisement call, while type II males sneak spawn to steal fertilizations from type I males. Like females, type II males can only produce brief, agonistic, grunt type vocalizations. Here, we tested the hypothesis that intrasexual differences in the number of CA neurons and their fiber innervation patterns throughout the vocal motor pathway may provide neural substrates underlying divergence in reproductive behavior between morphs. We employed immunofluorescence (-ir) histochemistry to measure tyrosine hydroxylase (TH; a rate-limiting enzyme in catecholamine synthesis) neuron numbers in several forebrain and hindbrain nuclei as well as TH-ir fiber innervation throughout the vocal pathway in type I and type II males collected from nests during the summer reproductive season. After controlling for differences in body size, only one group of CA neurons displayed an unequivocal difference between male morphs: the extraventricular vagal-associated TH-ir neurons, located just lateral to the dimorphic vocal motor nucleus (VMN), were significantly greater in number in type II males. In addition, type II males exhibited greater TH-ir fiber density within the VMN and greater numbers of TH-ir varicosities with putative contacts on vocal motor neurons. This strong inverse relationship between the predominant vocal morphotype and the CA innervation of vocal motor neurons suggests that catecholamines may function to inhibit vocal output in midshipman. These findings support catecholamines as direct modulators of vocal behavior, and differential CA input appears reflective of social and reproductive behavioral divergence between male midshipman morphs.
© 2015 S. Karger AG, Basel.
Figures
Similar articles
-
Sexually-dimorphic expression of tyrosine hydroxylase immunoreactivity in the brain of a vocal teleost fish (Porichthys notatus).J Chem Neuroanat. 2014 Mar;56:13-34. doi: 10.1016/j.jchemneu.2014.01.001. Epub 2014 Jan 10. J Chem Neuroanat. 2014. PMID: 24418093
-
Exposure to advertisement calls of reproductive competitors activates vocal-acoustic and catecholaminergic neurons in the plainfin midshipman fish, Porichthys notatus.PLoS One. 2013 Aug 6;8(8):e70474. doi: 10.1371/journal.pone.0070474. Print 2013. PLoS One. 2013. PMID: 23936438 Free PMC article.
-
Catecholaminergic innervation of central and peripheral auditory circuitry varies with reproductive state in female midshipman fish, Porichthys notatus.PLoS One. 2015 Apr 7;10(4):e0121914. doi: 10.1371/journal.pone.0121914. eCollection 2015. PLoS One. 2015. PMID: 25849450 Free PMC article.
-
A tale of two males: Behavioral and neural mechanisms of alternative reproductive tactics in midshipman fish.Horm Behav. 2024 May;161:105507. doi: 10.1016/j.yhbeh.2024.105507. Epub 2024 Mar 12. Horm Behav. 2024. PMID: 38479349 Review.
-
Adaptive hearing in the vocal plainfin midshipman fish: getting in tune for the breeding season and implications for acoustic communication.Integr Zool. 2009 Mar;4(1):33-42. doi: 10.1111/j.1749-4877.2008.00133.x. Integr Zool. 2009. PMID: 21392275 Review.
Cited by
-
Inhibitory and modulatory inputs to the vocal central pattern generator of a teleost fish.J Comp Neurol. 2018 Jun 1;526(8):1368-1388. doi: 10.1002/cne.24411. Epub 2018 Feb 28. J Comp Neurol. 2018. PMID: 29424431 Free PMC article.
-
Attention and Motivated Response to Simulated Male Advertisement Call Activates Forebrain Dopaminergic and Social Decision-Making Network Nuclei in Female Midshipman Fish.Integr Comp Biol. 2017 Oct 1;57(4):820-834. doi: 10.1093/icb/icx053. Integr Comp Biol. 2017. PMID: 28992072 Free PMC article.
References
-
- Appeltants D, Ball GF, Balthazart J. The distribution of tyrosine hydroxylase in the canary brain: demonstration of a specific and sexually dimorphic catecholaminergic innervation of the telencephalic song control nulcei. Cell Tissue Res. 2001;304:237–259. - PubMed
-
- Appeltants D, Ball GF, Balthazart J. The origin of catecholaminergic inputs to the song control nucleus RA in canaries. Neuroreport. 2002a;13:649–653. - PubMed
-
- Appeltants D, Ball GF, Balthazart J. Noradrenergic control of auditory information processing in female canaries. Behav Brain Res. 2002b;133:221–235. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
