

Stable 5-Hydroxymethylcytosine (5hmC) Acquisition Marks Gene Activation During Chondrogenic Differentiation
- PMID: 26363184
- PMCID: PMC4860191
- DOI: 10.1002/jbmr.2711
Stable 5-Hydroxymethylcytosine (5hmC) Acquisition Marks Gene Activation During Chondrogenic Differentiation
Abstract
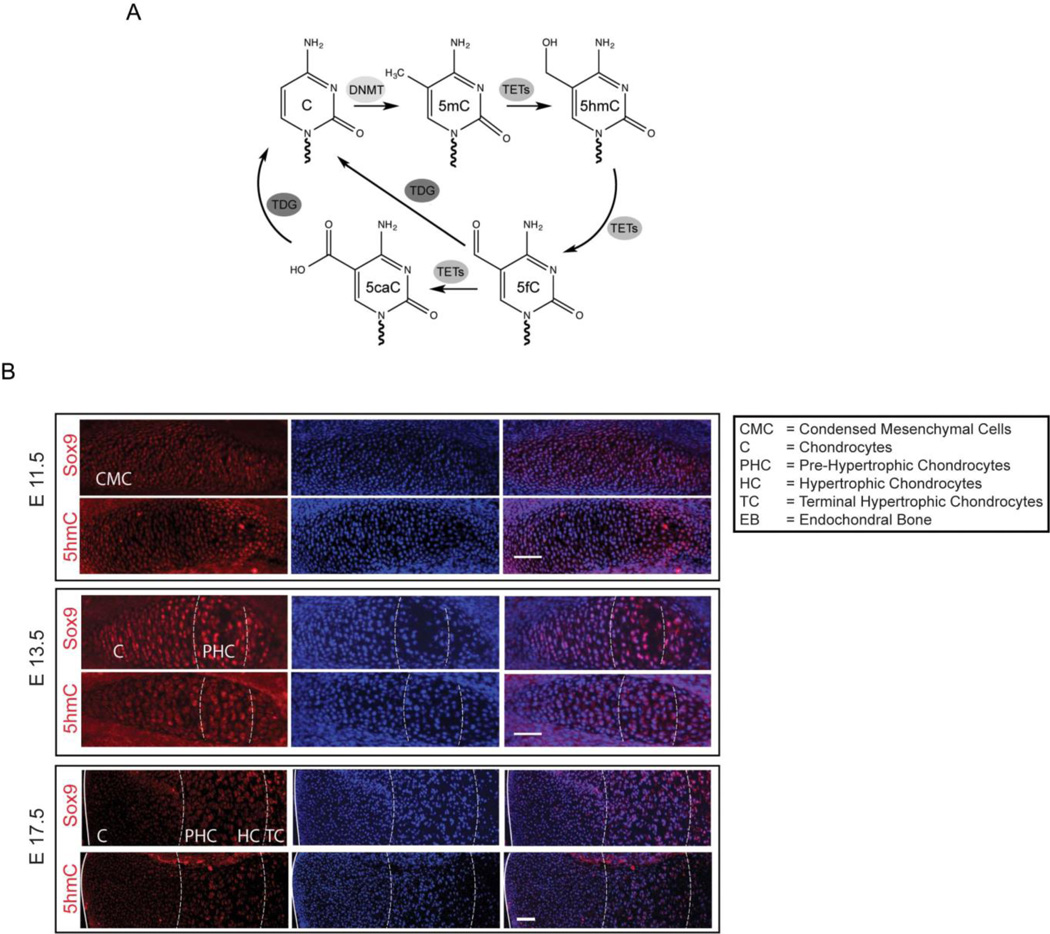
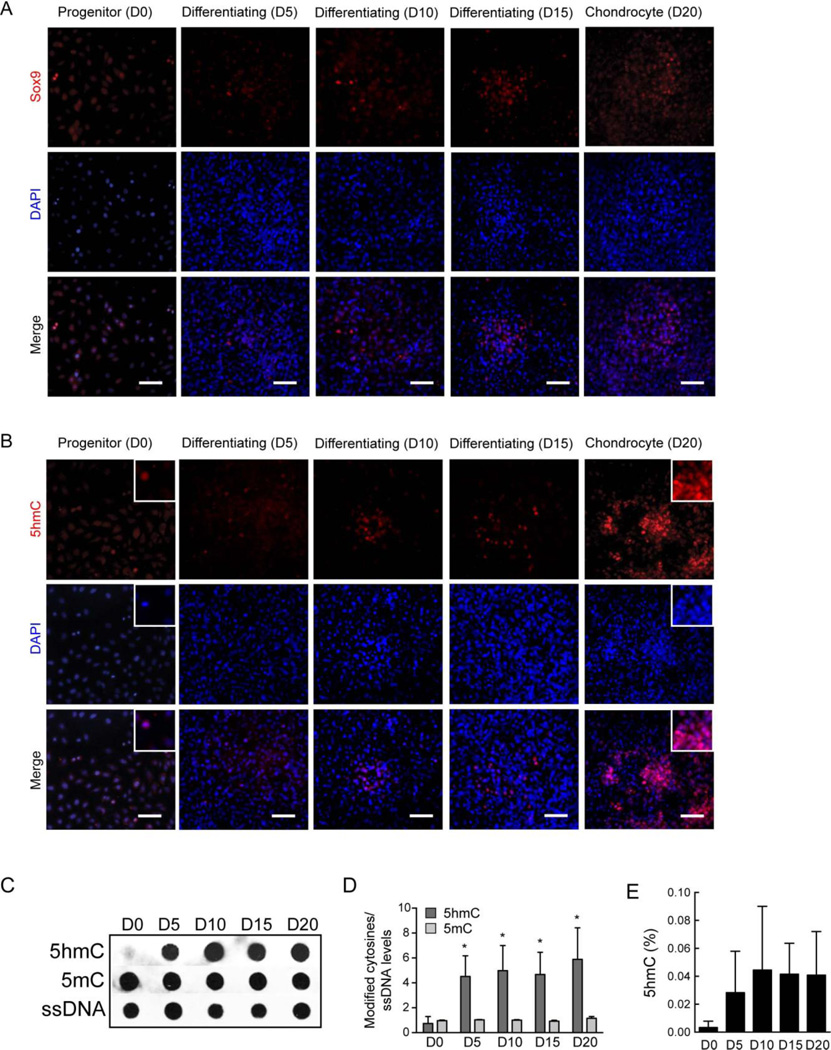
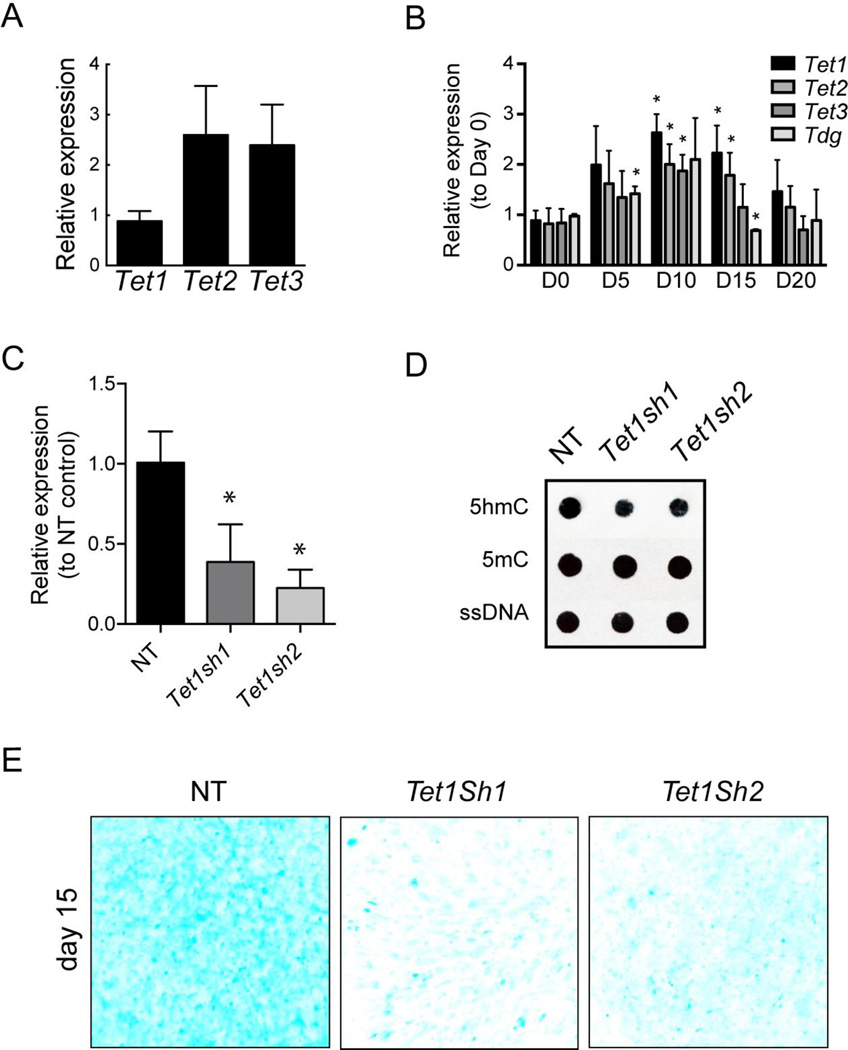
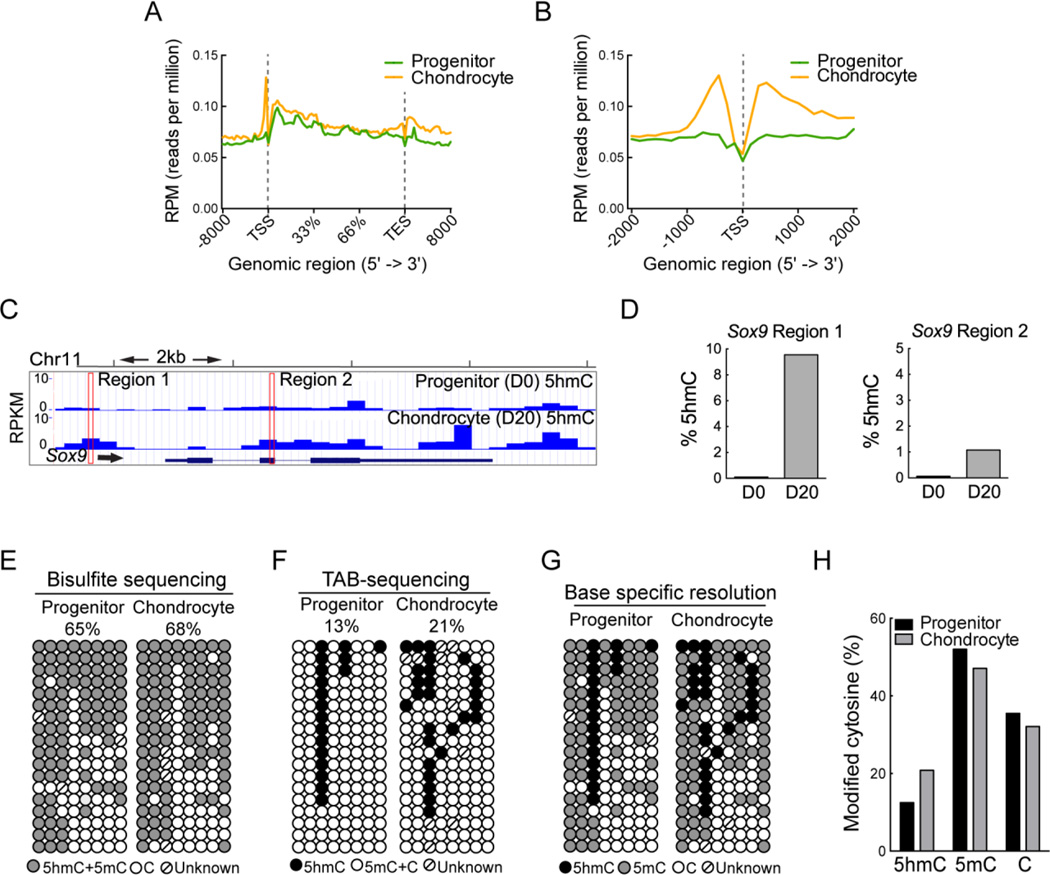
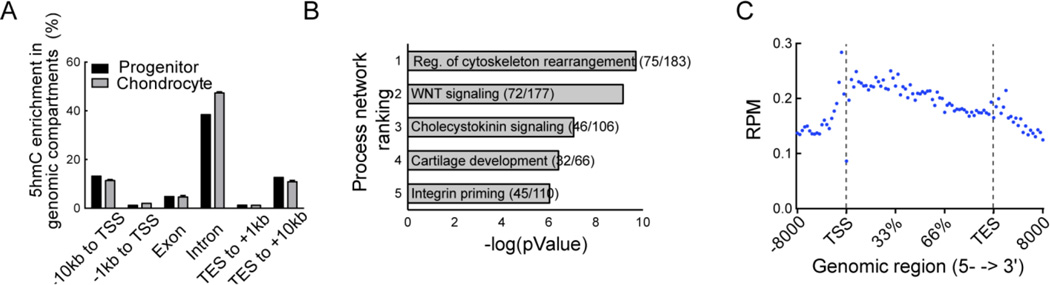
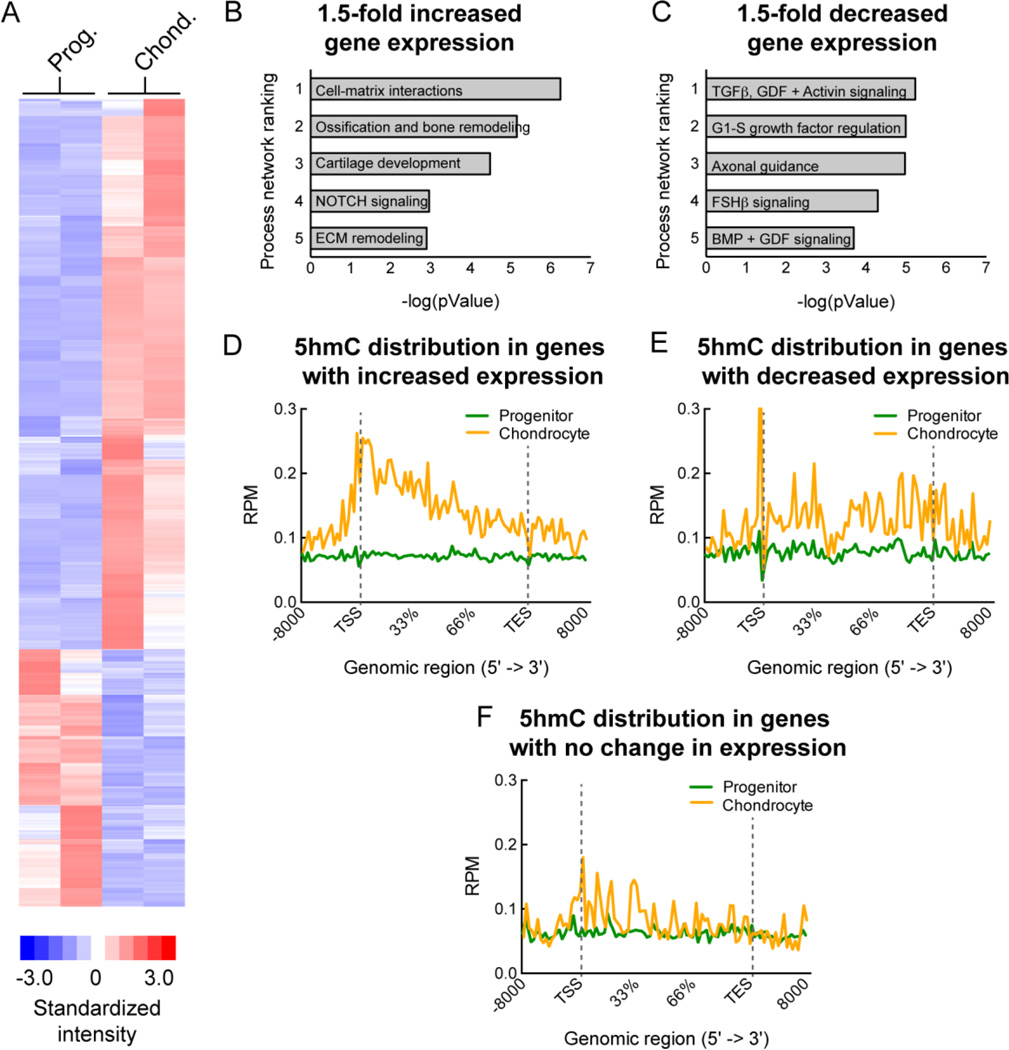
Regulation of gene expression changes during chondrogenic differentiation by DNA methylation and demethylation is little understood. Methylated cytosines (5mC) are oxidized by the ten-eleven-translocation (TET) proteins to 5-hydroxymethylcytosines (5hmC), 5-formylcytosines (5fC), and 5-carboxylcytosines (5caC), eventually leading to a replacement by unmethylated cytosines (C), ie, DNA demethylation. Additionally, 5hmC is stable and acts as an epigenetic mark by itself. Here, we report that global changes in 5hmC mark chondrogenic differentiation in vivo and in vitro. Tibia anlagen and growth plate analyses during limb development at mouse embryonic days E 11.5, 13.5, and 17.5 showed dynamic changes in 5hmC levels in the differentiating chondrocytes. A similar increase in 5hmC levels was observed in the ATDC5 chondroprogenitor cell line accompanied by increased expression of the TET proteins during in vitro differentiation. Loss of TET1 in ATDC5 decreased 5hmC levels and impaired differentiation, demonstrating a functional role for TET1-mediated 5hmC dynamics in chondrogenic differentiation. Global analyses of the 5hmC-enriched sequences during early and late chondrogenic differentiation identified 5hmC distribution to be enriched in the regulatory regions of genes preceding the transcription start site (TSS), as well as in the gene bodies. Stable gains in 5hmC were observed in specific subsets of genes, including genes associated with cartilage development and in chondrogenic lineage-specific genes. 5hmC gains in regulatory promoter and enhancer regions as well as in gene bodies were strongly associated with activated but not repressed genes, indicating a potential regulatory role for DNA hydroxymethylation in chondrogenic gene expression.
Keywords: 5-HYDROXYMETHYLCYTOSINE; CHONDROGENESIS; DIFFERENTIATION; DNA DEMETHYLATION; EPIGENETICS.
© 2015 American Society for Bone and Mineral Research.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
