

Structural Basis of Telomerase Inhibition by the Highly Specific BIBR1532
- PMID: 26365799
- PMCID: PMC4598299
- DOI: 10.1016/j.str.2015.08.006
Structural Basis of Telomerase Inhibition by the Highly Specific BIBR1532
Abstract
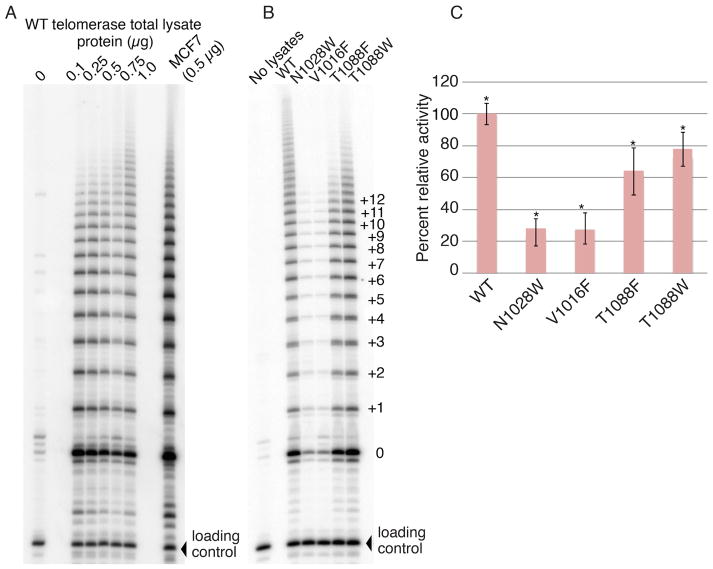
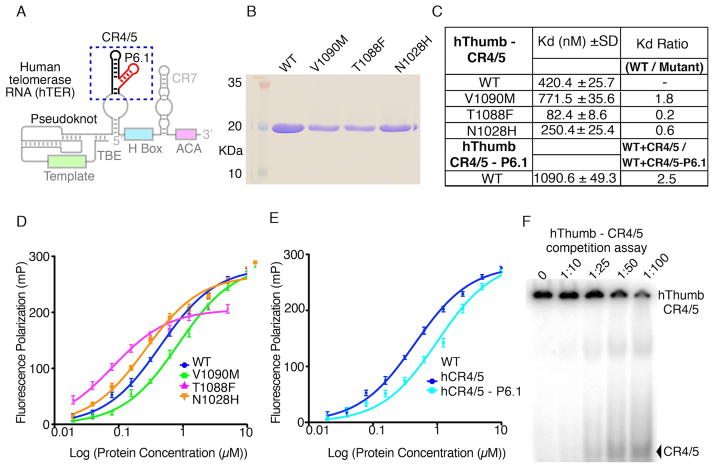
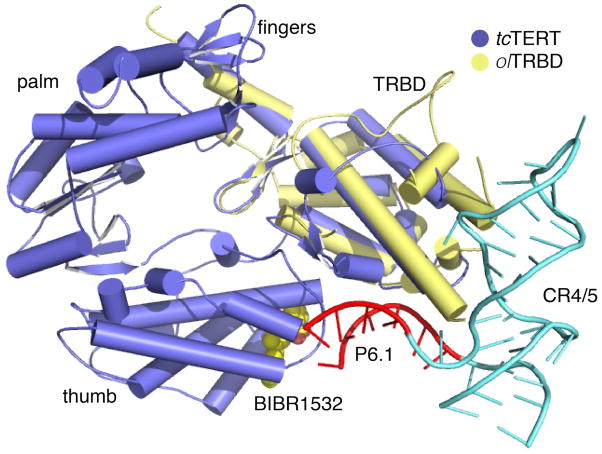
BIBR1532 is a highly specific telomerase inhibitor, although the molecular basis for inhibition is unknown. Here we present the crystal structure of BIBR1532 bound to Tribolium castaneum catalytic subunit of telomerase (tcTERT). BIBR1532 binds to a conserved hydrophobic pocket (FVYL motif) on the outer surface of the thumb domain. The FVYL motif is near TRBD residues that bind the activation domain (CR4/5) of hTER. RNA binding assays show that the human TERT (hTERT) thumb domain binds the P6.1 stem loop of CR4/5 in vitro. hTERT mutations of the FVYL pocket alter wild-type CR4/5 binding and cause telomere attrition in cells. Furthermore, the hTERT FVYL mutations V1025F, N1028H, and V1090M are implicated in dyskeratosis congenita and aplastic anemia, further supporting the biological and clinical relevance of this novel motif. We propose that CR4/5 contacts with the telomerase thumb domain contribute to telomerase ribonucleoprotein assembly and promote enzymatic activity.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
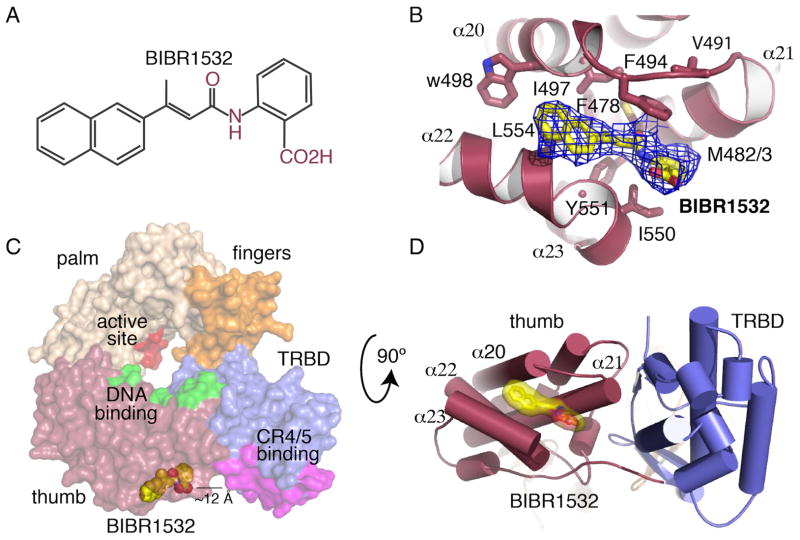
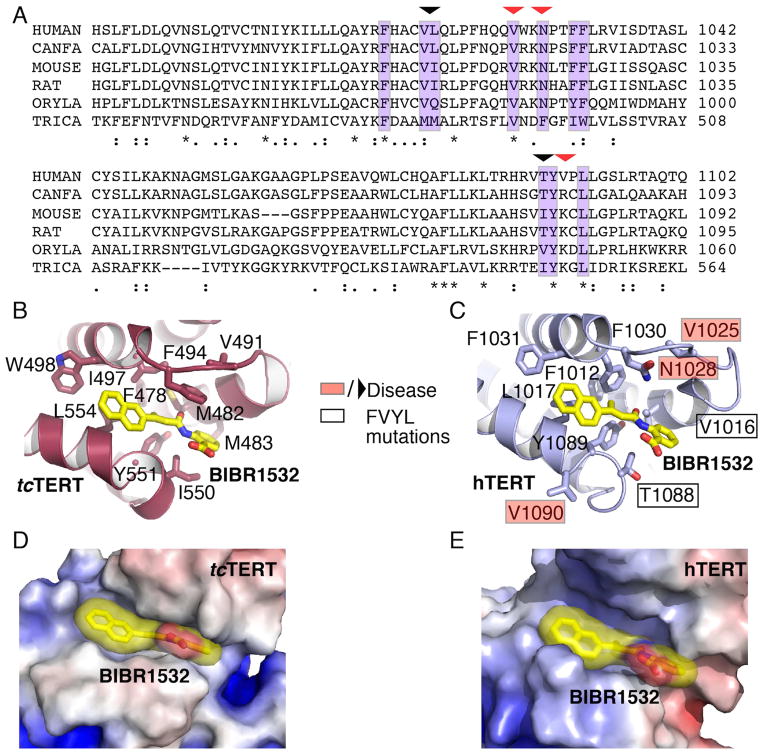
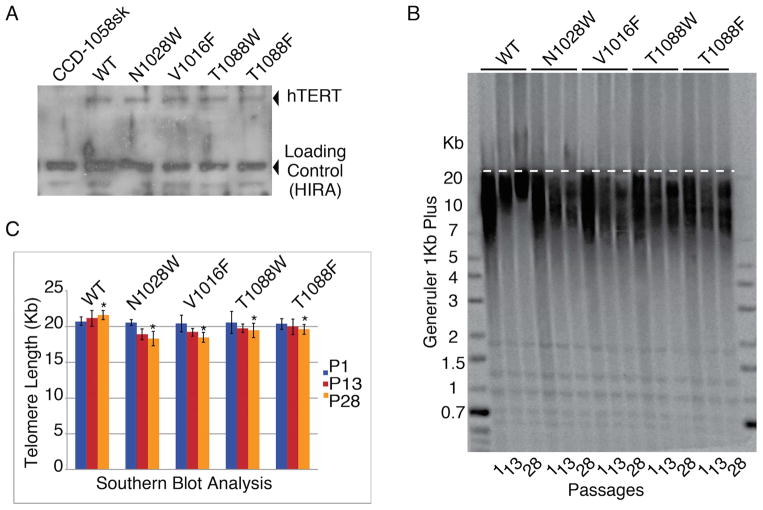
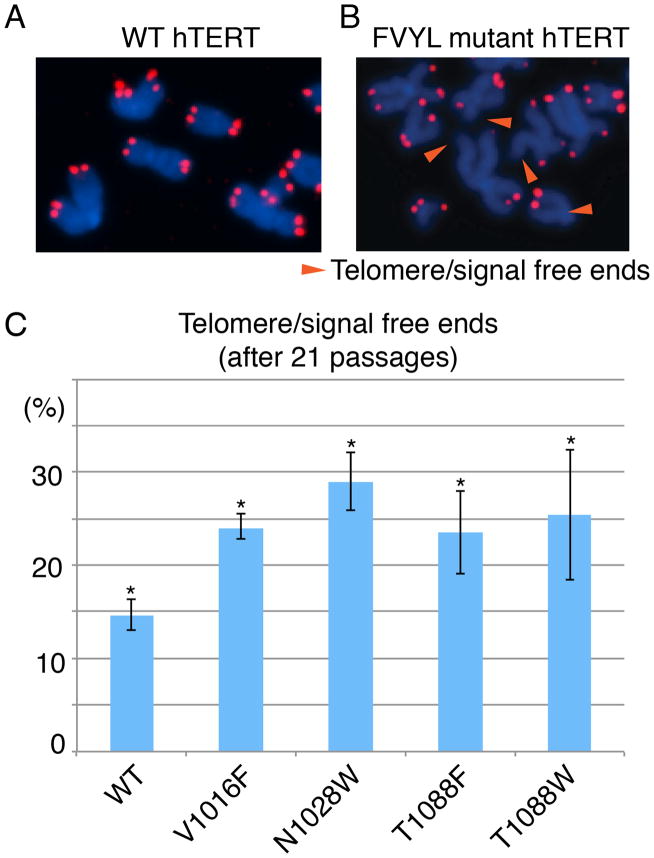
References
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 1976;72:248–254. - PubMed
-
- Damm K, Hemmann U, Garin-Chesa P, Hauel N, Kauffmann I, Priepke H, Niestroj C, Daiber C, Enenkel B, Guilliard B, Lauritsch I, Muller E, Pascolo E, Sauter G, Pantic M, Martens UM, Wenz C, Lingner J, Kraut N, Rettig WJ, Schnapp A. A highly selective telomerase inhibitor limiting human cancer cell proliferation. The EMBO journal. 2001;20:6958–6968. - PMC - PubMed
-
- El-Daly H, Kull M, Zimmermann S, Pantic M, Waller CF, Martens UM. Selective cytotoxicity and telomere damage in leukemia cells using the telomerase inhibitor BIBR1532. Blood. 2005;105:1742–1749. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
