

Pycnogenol Attenuates the Release of Proinflammatory Cytokines and Expression of Perilipin 2 in Lipopolysaccharide-Stimulated Microglia in Part via Inhibition of NF-κB and AP-1 Activation
- PMID: 26367267
- PMCID: PMC4569068
- DOI: 10.1371/journal.pone.0137837
Pycnogenol Attenuates the Release of Proinflammatory Cytokines and Expression of Perilipin 2 in Lipopolysaccharide-Stimulated Microglia in Part via Inhibition of NF-κB and AP-1 Activation
Abstract
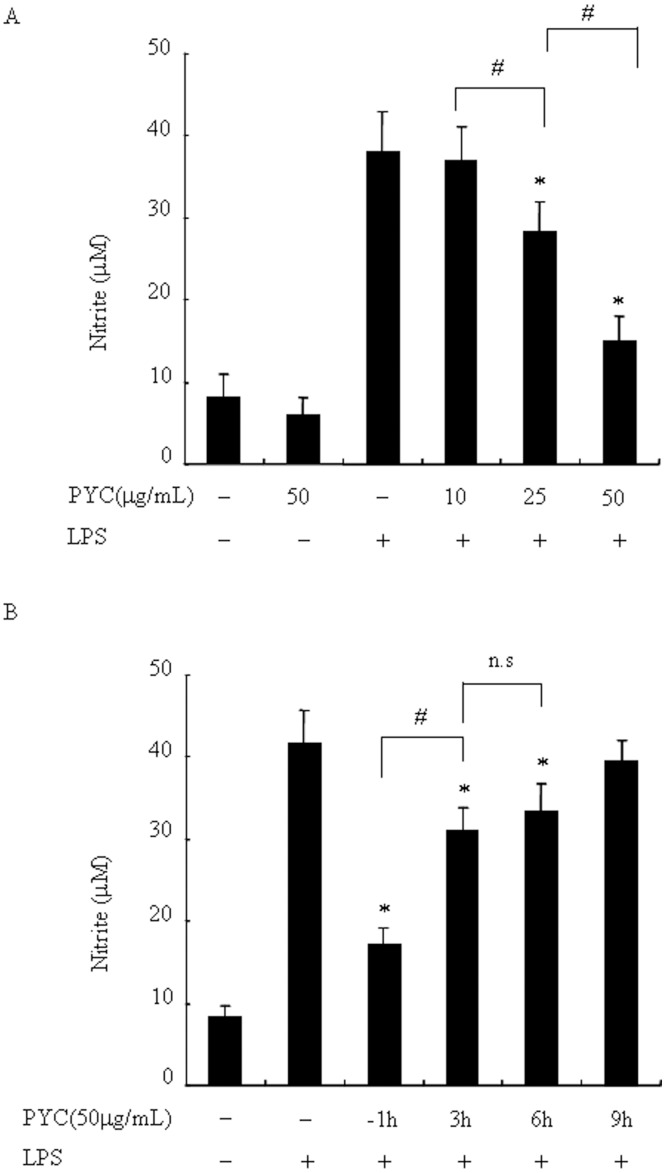
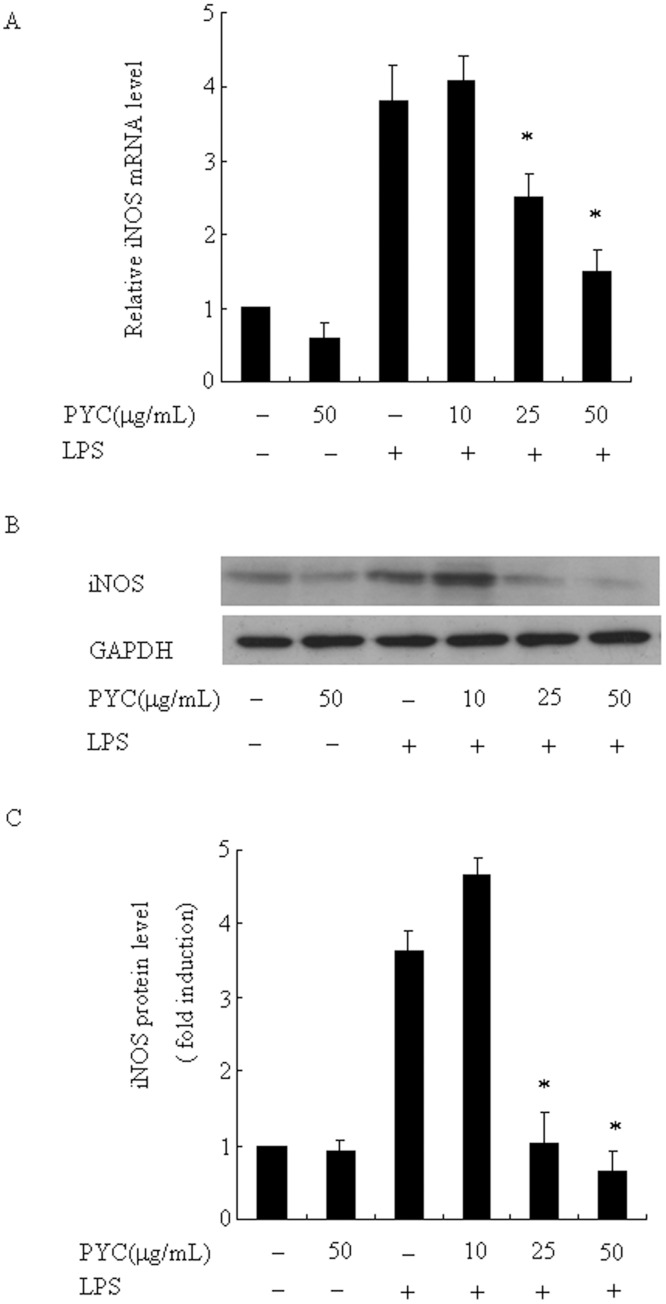
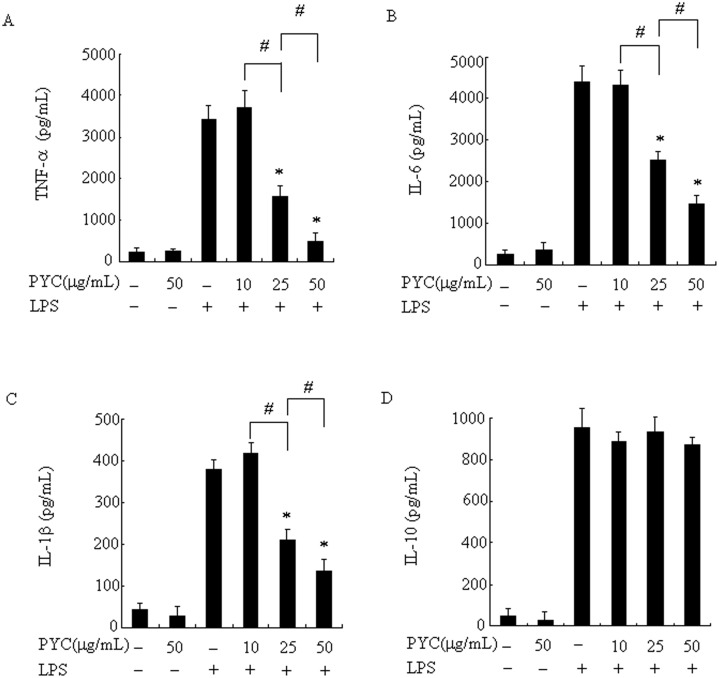
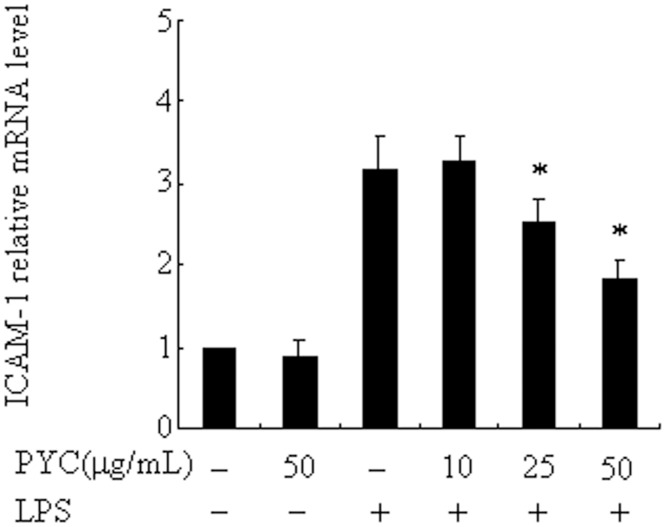
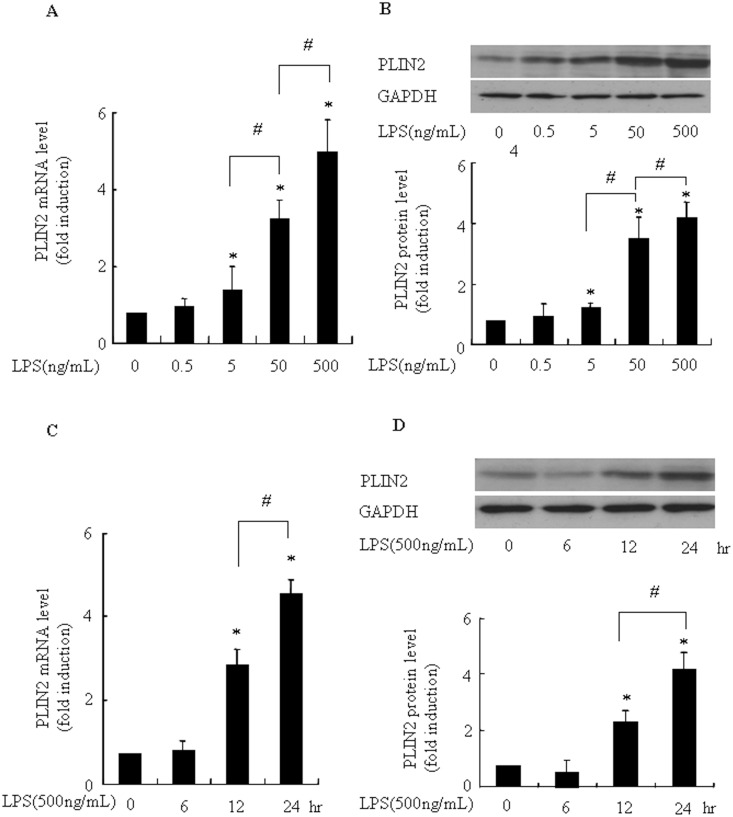
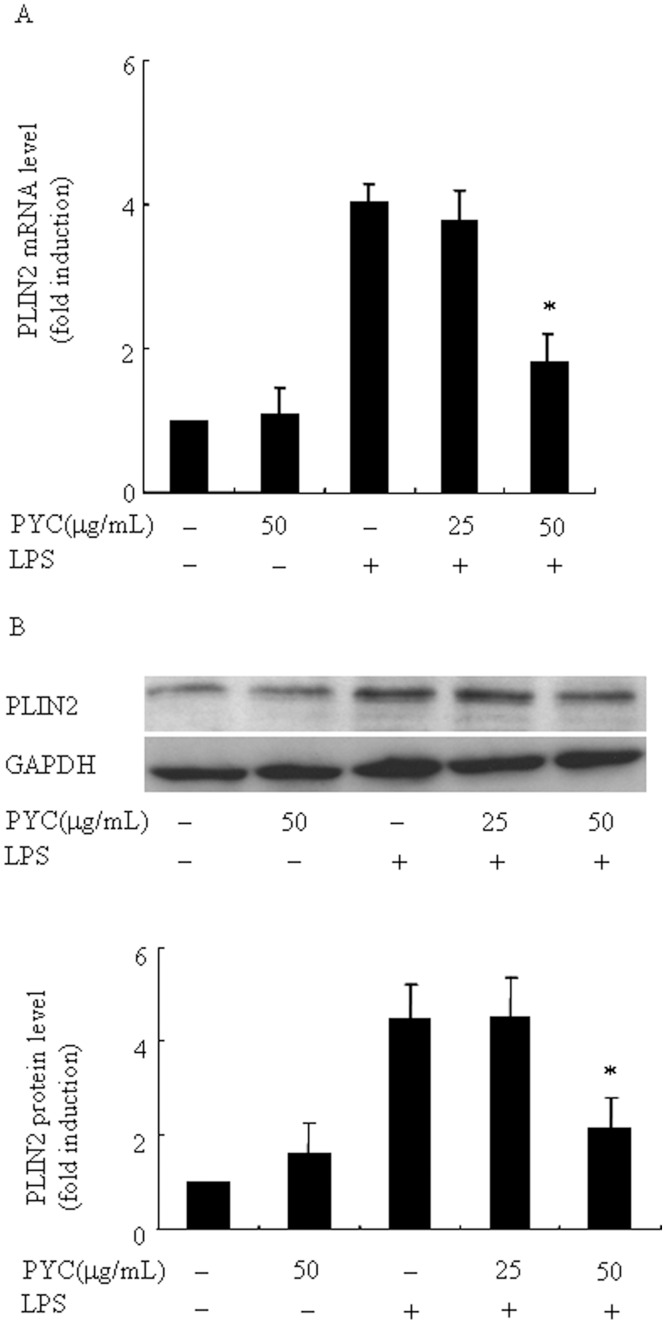
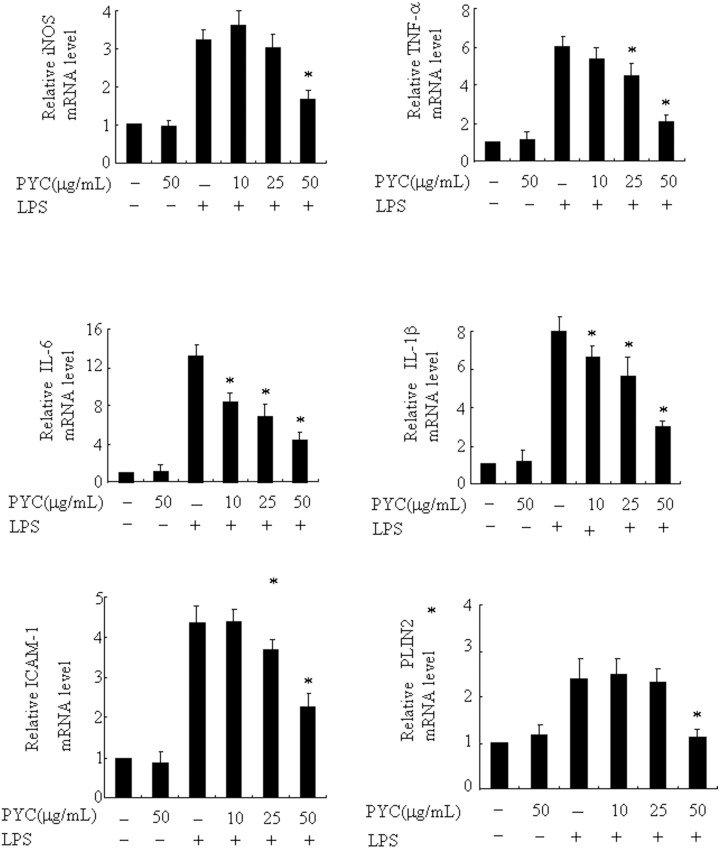
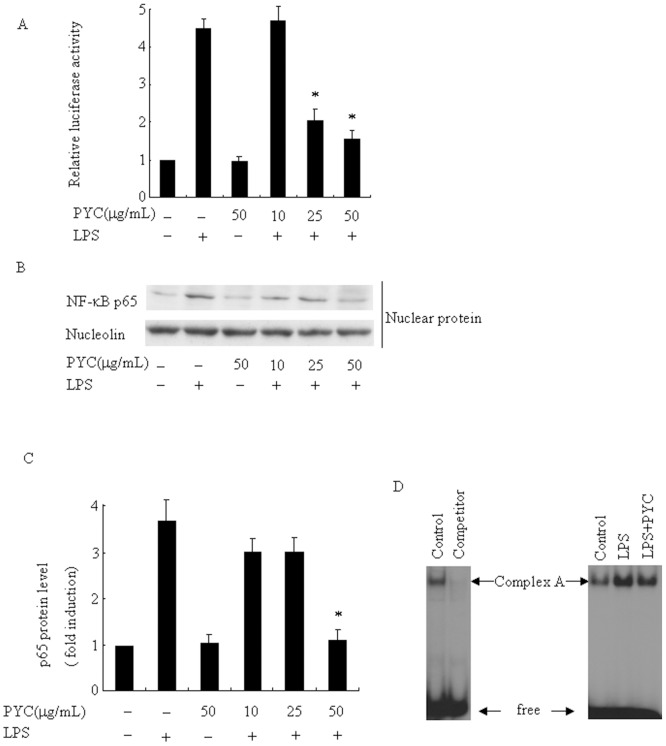
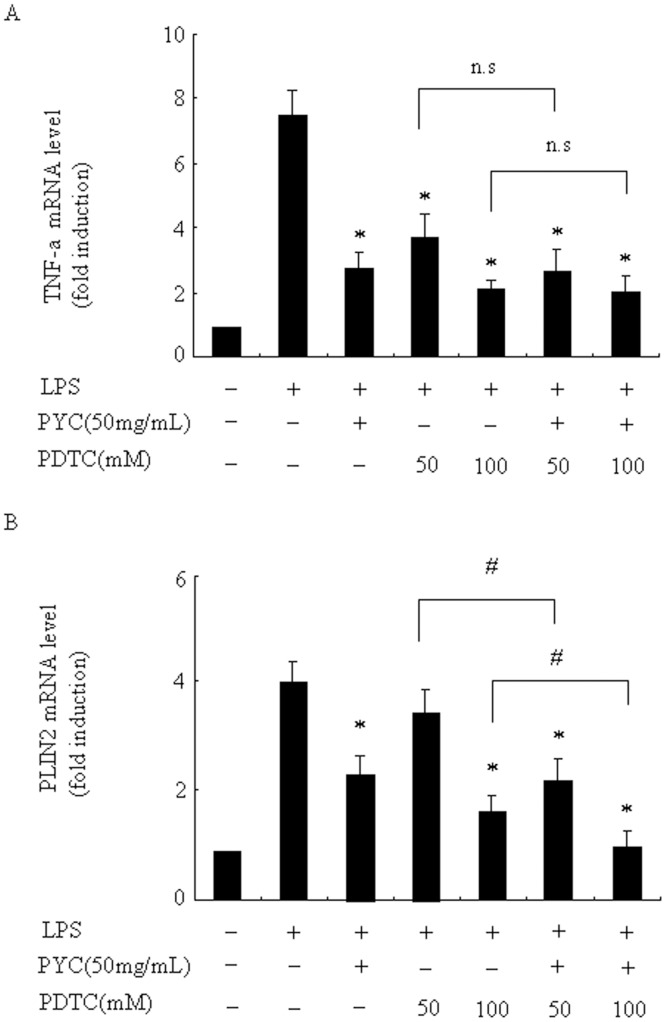
Over activation of microglia results in the production of proinflammatory agents that have been implicated in various brain diseases. Pycnogenol is a patented extract from French maritime pine bark (Pinus pinaster Aiton) with strong antioxidant and anti-inflammatory potency. The present study investigated whether pycnogenol may be associated with the production of proinflammatory mediators in lipopolysaccharide-stimulated BV2 (mouse-derived) microglia. It was found that pycnogenol treatment was dose-dependently associated with significantly less release of nitricoxide (NO), TNF-α, IL-6 and IL-1β, and lower levels of intercellular adhesion molecule1 (ICAM-1) and perilipin 2 (PLIN2). Furthermore, this effect was replicated in primary brain microglia. Levels of inducible NO synthase mRNA and protein were attenuated, whereas there was no change in the production of the anti-inflammatory cytokine IL-10. Further evidence indicated that pycnogenol treatment led to the suppression of NF-κB activation through inhibition of p65 translocation into the nucleus and inhibited DNA binding of AP-1, suggesting that these proinflammatory factors are associated with NF-κB and AP-1. We conclude that pycnogenol exerts anti-inflammatory effects through inhibition of the NF-κB and AP-1pathway, and may be useful as a therapeutic agent in the prevention of diseases caused by over activation of microglia.
Conflict of interest statement
Figures

References
-
- Giulian D, Li J, Li X, George J, Rutecki PA. The impact of microglia-derived cytokines upon gliosis in the CNS. Dev Neurosci. 1994;16(3–4):128–36. Epub 1994/01/01. . - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
